The species-rich phylum Microsporidia includes obligate intracellular parasites that infect a wide range of vertebrate and invertebrate hosts. Microsporidians have evolved an elaborate mechanism for invading animal host cells, but have otherwise greatly reduced biological complexity. The smallest known autonomous nuclear genome is that of a microsporidian, Encephalitozoon intestinalis (Keeling and Corradi 2011). Although the taxonomic affiliation of the Microsporidia has long been controversial, they are now known to either fall within the Fungi or to be extremely closely related to the Fungi.
Microsporidians produce resistant spores and possess a unique organelle, the polar tubule, or polar filament, which is coiled inside the spore (see below). The spores of microsporidians known to infect humans range from 1 to 4 microns in diameter. At least 14 microsporidian species have been identified as human pathogens (see Table from U.S. CDC): Brachiola algerae, B. connori, B. vesicularum, Encephalitozoon cuniculi, Enc. hellem, Enc. intestinalis (formerly known as Septata intestinalis), Enterocytozoon bieneusi, Microsporidium ceylonensis, M. africanum, Nosema ocularum, Pleistophora sp., Trachipleistophora hominis, T. anthropophthera, and Vittaforma corneae. Some domestic and wild animals may be naturally infected with Enc. cuniculi, Enc. intestinalis, and Ent. bieneusi (e.g., Santin and Fayer 2011). Birds, especially parrots (parakeets, lovebirds, budgies) are naturally infected with Enc. hellem. Enterocytozoon bieneusi and V. corneae have been identified in surface waters and spores of "Nosema" (probably B. algerae) have been identified in ditch water.
The infective form of microsporidians is the resistant spore, which can survive for long periods in the environment. The spore extrudes its polar tubule and infects the host cell. The spore injects the infective sporoplasm into the eukaryotic host cell through the polar tubule. Inside the cell, the sporoplasm undergoes extensive multiplication either by merogony (binary fission) or schizogony (multiple fission). This development can occur either in direct contact with the host cell cytoplasm (e.g., Enterocytozoon bieneusi) or inside a vacuole known as a parasitophorous vacuole (e.g., Encephalitozoon intestinalis). Either free in the cytoplasm or inside a parasitophorous vacuole, microsporidia develop by sporogony to mature spores. During sporogony, a thick wall is formed around the spore, which provides resistance to adverse environmental conditions. When the spores increase in number and completely fill the host cell cytoplasm, the cell membrane is disrupted and releases the spores to the surroundings. These free mature spores can infect new cells, continuing the cycle. (Centers for Disease Control Parasites and Health Website)
Microsporidia are being increasingly recognized as opportunistic infectious agents worldwide (Didier and Weiss 2011). Cases of microsporidiosis have been reported from developed as well as developing countries, including Argentina, Australia, Botswana, Brazil, Canada, Czech Republic, France, Germany, India, Italy, Japan, The Netherlands, New Zealand, Spain, Sri Lanka, Sweden, Switzerland, Thailand, Uganda, United Kingdom, United States of America, and Zambia. (Centers for Disease Control Parasites and Health Website)
Nosema microsporidians have been suspected of playing an important role in the dramatic population declines of certain bumblebee species (e.g., Cameron et al. 2011 and references therein, but see Kissinger et al. 2011) and some researchers believe Nosema has played a role in honeybee colony collapse disorder in at least some regions (Paxton 2010).
Vertical transmission (i.e., from parent host to offspring) is unusually common among microsporidians. Because this mode of transmission depends on a host surviving long enough to reproduce, it is often associated with low pathogenesis. Smith (2009) reviewed the ecology and evolution of microsporidians, emphasizing both what is known and what is not known.
Los microsporidios (Microsporidia) son un conxuntu de microorganismos parásitos intracelulares d'animales. Hai descrites unes 800 especies.
Fueron clasificaos como protozóos, como protistes y tamién como fungos. En realidá son un grupu basal d'fungos parásitos obligaos que baxen d'organismos unicelulares que nel procesu evolutivu de la so adaptación a la vida parasitaria, perdieron les mitocondrias.
Los microsporidios parasitan les célules intestinales de pexes y aves, anque tamién s'atoparon n'invertebraos (Nosema produz la pebrina, una enfermedá de los viermes de seda Bombyx mori, como describió Luis Pasteur).
Nosema apis ye un microsporidiu que causa una enfermedá denominada nosemosis, qu'ataca a l'abeya melífera, causándo-y inflamación intestinal y foria. L'antibióticu utilizáu nel tratamientu d'esta enfermedá ye la fumagilina.
Los microsporidios (Microsporidia) son un conxuntu de microorganismos parásitos intracelulares d'animales. Hai descrites unes 800 especies.
Fueron clasificaos como protozóos, como protistes y tamién como fungos. En realidá son un grupu basal d'fungos parásitos obligaos que baxen d'organismos unicelulares que nel procesu evolutivu de la so adaptación a la vida parasitaria, perdieron les mitocondrias.
Bu tipin nümayəndələri həm onurğalıların, həm də onurğasızların orqanizmlərində parazitlik edir. 1500-dən çox növü məlum olan mikrosporidilər hüceyrədaxili parazit həyat sürür. Ən geniş yayılmış mikrosporidilər bal arılarında ağ diareya yaradan Nosema apis və Nosema bombycis-dir.
Spor daxilində bir iki ədəd rüşeym və unikal orqanel kompleksi olan ekstruziya aparatı yerləşir. Bu aparat sahibin hüceyrəsinin xarici qişasını dəlib onu yoluxdurmaq, rüşeymləri bilavasitə sitoplazmaya yeritmək üçündür. Hüceyrədə mikrosporidi proliferativ siklə (meroqon) məruz qalır. Sonda spor yaranır (sporoqon).[1]
ilk öncə balıq parazitin sporlarını udur. İlk infeksiya zədələnməsi mədə-bağırsaq traktının yuxarı hissəsinə təsadüf edir. Ona görə də ilk ksenomlar bağırsağın ilk şöbələrinə təsadüf edir. Parazitin sonrakı yayılması (böyrəklər, dalaq, yumurtalıq, göz, dərialtı birləşdirici toxumalar, üzmə qovuqcuğunun divarları və s.) immun sisteminin hüceyrələri vasitəsilə həyata keçir. Ksemonlar məhz bu yerlərdə yaranır.
Ksenom – parazitin təsirilə sahib orqanizminin daxilində baş vermiş proliferasiya nəticəsində öz strukturunu tamamilə dəyişmiş sahib orqanizminin hüceyrəsidir. Bu zaman hər iki komponent morfoloji və fizioloji cəhətdən sıx birləşərək sahibin orqanizmində müstəqil inkişaf edən unikal vəhdət yaradırlar. Makroskopik ksenomlar uzunluğu 1 mm-ə çatan ağ oval düyünlərə bənzəyib düyün topaları yaradır.
Bəzi mikrosporidilər (Multivalvulida, Bivalvulida) balıqların təhlükəli xəstəlik törədiciləridir. Balıqlar 150-dən çox mikrosporidinin (14 cins) sabibi qismində çıxış edir. Hüceyrədaxili obliqat parazitliyi şərtləndirən əsas səbəb onlarda mitoxondrinin və digər orqanellərin olmamasıdır. Ona görə də mikrosporidilər sahib hüceyrəsindən kənarda yalnız spor şəklində ola bilir. uğumayaqlılar və balıqlar arasında ən geniş yayılmış mikrosporidilərdən biri də Glugea anomala və Nucleospora secunda parazitləridir.[2]
Glugea anomala (Moniez, 1887) üçiynəli (Gasterosteus aculeatus) və doqquziynəli tikanbalıqlarının (Pungitius pungitius) parazitləridir. Onlar həmçinin Nothobranchius eggersi, N. korthausae kimi notobranxiusları və s. balıq növlərini, bir sıra akvarium balıqlarını zədələyir.
İlk öncə balıq parazitin sporlarını udur. İlk infeksiya zədələnməsi mədə-bağırsaq traktının yuxarı hissəsinə təsadüf edir. Ona görə də ilk ksenomlar bağırsağın ilk şöbələrinə təsadüf edir. Parazitin sonrakı yayılması (böyrəklər, dalaq, yumurtalıq, göz, dərialtı birləşdirici toxumalar, üzmə qovuqcuğunun divarları və s.) immun sisteminin hüceyrələri vasitəsilə həyata keçir. Ksemonlar məhz bu yerlərdə yaranır. Mikrosporidilər ipəkqurdu və arı təsərrüfatlarına ciddi ziyan vurur. Belə ki onlar bal arılarında (Apis mellifera) və tut ipəkqurdunda (Bombyx mori) nozematoz epizootiyası yaradır, midiya (Mytilus edulus) və s. kimi bir sıra faydalı onurğasızların və bir sıra sənaye əhəmiyyətli balıqların kütləvi məhvinə səbəb olur.
İnsan mikrosporidiozu xüsusilə təhlükəlidir. Adətən o, immun çatışmazlığı fonunda ağır və sürətlə inkişaf edən opportunistik xəstəliklər yaradır.Encephalitozoon, Enterocytozoon, Nosema, Pleistophora, Trachipleistophora, Vittaforma və s. nümayəndələr insan üçün patogen sayılan mikrosporidilərdir. VİÇ-infeksiyası dövrü Enterocytozoon bieneusi, Encephalitozoonintestinalis, Encephalitozoon hellem, Trachipleistophora hominis və Trachipleistophora anthropophtera kimi insan mikrosporidiozu törədiciləri təsvir edilmişdir. Sübut edilmişdir ki, dovşanlarda parazitlik edən Encephalitozoon cuniculimikrosporidisi normal immunkompetent insanları yoluxdurur. Yoluxmanın ən geniş yayılmış simptomu diareyadır.
Zərərverici həşəratlarda parazitlik edən mikrosporidilər kənd təsərrüfatı, tibb və veterianriyada effektiv bioloji mübarizə agentləridir. Bəzi ağcaqanad mikrosporidilərinin növləri onların sürfələrini öldürür, dişi fərdlərin nəslverməsini kəskin azaldır, malyariya plazmodilərinin inkişafını dayandırır. İlkin fazadaT. hominissporu öz daxilini zülal quruluşlu qütb borucuğu vasitəsilə sahib hüceyrəyə yeridir (inyeksiya edir). Bu birnüvəli sporoplazm sahib hüceyrənin yoluxmasının ilkin fazası hesab edilir. İnyeksiyadan sonra parazit (meront) böyüyür və sahib hüceyrənin daxilində proliferasiya edir. Meroqoniya parazitin həyat siklinin ikinci fazası sayılır. Sporların formalaşması (sporoqoniya) zamanı mikrosporidilərin sporlarının divarları yığılır və T. hominisproliferasiyanınnövbəti mərhələsinə keçir. Sonda yetkin sporlar sahib hüceyrənin daxilində formalaşır (5-ci mərhələ). Bu sporlar öz növbəsində öz qütb borucuqlarını sahib hüceyrədən çıxarır və qonşu hüceyrəni yoluxdurur.
Xəzər dənizində Glugea, Pleistephora cinslərinə aid 8 ixtioparazit mikrosporidia növü təyin edilmişdir İbrahimov Ş., 2012. Yeni təsnifatlarda bu tip göbələklər aləminə aid edilir.
Bu tipin nümayəndələri həm onurğalıların, həm də onurğasızların orqanizmlərində parazitlik edir. 1500-dən çox növü məlum olan mikrosporidilər hüceyrədaxili parazit həyat sürür. Ən geniş yayılmış mikrosporidilər bal arılarında ağ diareya yaradan Nosema apis və Nosema bombycis-dir.
Els microsporidis (microsporidia) són un tipus de fongs extremadament minúsculs que són paràsits d'animals, sobretot insectes, però també crustacis, peixos o, fins i tot, humans; provocant, llavors, una microsporidiosi.
Mikrosporidie (Microsporidia, Microspora, česky též hmyzomorky) je skupina jaderných organismů, dnes zpravidla považovaná za samostatný kmen hub.[1] Její zástupci jsou vnitrobuněční paraziti, většinou vegetující v cytoplazmě hostitelů. Známe více než 1200 druhů v 144 rodech.[2]
Dříve byly hmyzomorky považovány za součást říše prvoci (například ve skupinách Myxosporidia, Actinomyxidia, Haplosporidia, Sporozoa[3]), posléze byly řazeny do hypotetické skupiny Archezoa (dnes se neužívá), ale na základě porovnávání určitých molekulárních znaků se postupně dospělo k názoru, že jsou mikrosporidie řazeny mezi houby.[2][4][5] Molekulární analýzy zpočátku ukazovaly na umístění do oddělení spájivé houby (Zygomycota), dnes však dokládají příbuznost mikrosporidií s taxony Afelida a Rozellida/Cryptomycota, se kterými pravděpodobně tvoří přirozený „klad ARM“ (též zvaný Opisthosporidia) na bázi fylogenetického stromu hub (nebo jako jejich sesterskou skupinu v rámci nadřazených Holomycota).[6][7]
Mikrosporidie se vyznačují řadou znaků, které považujeme za primitivní a dříve byly důvodem řazení mezi prvoky. Mají totiž 70S ribozomy (podjednotky rRNA mají 16S a 23S). Chybí jim centrioly i jakékoliv respirační organely (hydrogenozomy, mitochondrie).[2]
Spory mají v klidové fázi jedno nebo několik jader, hrubé endoplazmatické retikulum, ribozomy, vakuolu a drobné váčky.[2]
Mnohé mikrosporidie hrají roli při vzniku onemocnění a mnohdy je dokonce vyvolávají, zejména u hmyzu. Jsou původci pébriny bource morušového, nosemózy včel, porcelánové nemoci raků, rybích infekcí, ale i člověka mohou napadnout, pokud má oslabený imunitní systém (např. u pacientů s onemocněním AIDS).[2]
Mikrosporidie (Microsporidia, Microspora, česky též hmyzomorky) je skupina jaderných organismů, dnes zpravidla považovaná za samostatný kmen hub. Její zástupci jsou vnitrobuněční paraziti, většinou vegetující v cytoplazmě hostitelů. Známe více než 1200 druhů v 144 rodech.
Die Microsporidia, auch Microspora oder Microsporea, sind einzellige, zu den Pilzen gehörende Parasiten und Erreger der Mikrosporidiasis. Sie können eine Größe von wenigen µm erreichen (2–12 µm). Microspora parasitieren meist intrazellulär bei Vertretern vieler Tierstämme, seltener in anderen Protisten aus der Sar genannten Klade der Eukaryoten. Eine Übertragung erfolgt durch Aufnahme einzelliger Sporen (meist oral, d. h. durch den Mund).
Mikrosporidien sind obligate Parasiten. Sie leben im Zellinneren einer Zelle ihres Wirts. Anders als viele andere Zellparasiten leben sie nicht in einer abgetrennten Vakuole, sondern direkt im Cytoplasma. Die Infektion erfolgt aus einer Spore, dem einzigen außerhalb der Wirtszelle vorkommenden Stadium. Das Cytoplasma (Sporoplasma genannt) des Parasiten ist von demjenigen der Wirtszelle durch seine Zellmembran getrennt, die oft noch eine Hülle vor allem aus komplexen Kohlenhydraten, Glycocalyx genannt, umgibt. Der Parasit schädigt in diesem Stadium die Wirtszelle zunächst nicht merklich (gelegentlich kommt es in Muskelfasern zu Funktionseinschränkungen), er vermehrt sich parallel zur Wirtszelle durch Zellteilung (Merogonie), das Stadium wird auch Meront genannt. Durch die Position innerhalb der Zelle ist er für das Immunsystem des Wirts unsichtbar. Die Parasitenzelle ist in ihrem Aufbau extrem reduziert und in basalen Zellfunktionen vollständig von der Wirtszelle abhängig. Sie enthält zunächst nur einen Zellkern (selten verdoppelt, Dikaryon), dicht gepackte Ribosomen und diverse, nicht zu Organellen organisierte Membranbestandteile, weder ein Golgi-Apparat noch funktionsfähige Mitochondrien sind erkennbar. Mitosomen genannte Organellen stellen reduzierte Relikte von Mitochondrien dar, in denen nur noch etwa 20 funktionale Proteine nachweisbar sind (normale Mitochondrien enthalten etwa 1000). Sie besitzen kein eigenständiges Genom mehr. Auch das Genom des Kerns der Mikrosporidien-Zelle ist hochgradig reduziert. Es sind nur noch etwa 2000 Gene vorhanden, so dass viele Stoffwechselprodukte aus der Wirtszelle übernommen werden müssen. So fehlen die Gene für die Nukleotid-Synthese und viele wesentliche Bestandteile der Atmungskette. Sie sind sogar teilweise auf den direkten Import von Adenosintriphosphat (ATP) aus der Wirtszelle zur Energieversorgung angewiesen, deren Stoffwechsel sie zu diesem Zweck anheizen. Obwohl auch die Ribosomen im Funktionsumfang reduziert sind, besitzt der Parasit extrem viele davon. Diese ermöglichen ihn ein sehr rasches Wachstum auf Kosten seines Wirts. Dessen Fähigkeit, sich gegen die Infektion durch programmierten Zelltod (Apoptose) zu wehren, wird durch Signalstoffe gezielt sabotiert. Im Ergebnis schwillt die infizierte Wirtszelle auf ein Vielfaches ihrer normalen Größe an. Einige Mikrosporidien (vor allem Fischparasiten) bilden im Inneren der, oft zu einem Syncytium verschmolzenen Wirtszellen riesige, mit Sporen gefüllte Einschlusskörper, die Xenosoma genannt werden.
Sehr schnell, oft innerhalb von drei Tagen nach der Infektion, beginnen die Mikrosporidien innerhalb der Wirtszelle mit der Produktion neuer Sporen. Das sporenbildende Stadium innerhalb der Zelle wird auch Sporont genannt. Bei den Gruppen, bei denen eine Meiose vorkommt, tritt diese unmittelbar vor der Sporenbildung auf. Die Sporen der Mikrosporidia dienen sowohl dazu, neue Zellen innerhalb des Wirts zu infizieren, als auch zur Infektion neuer Wirte. Für den Infektionsvorgang haben sie einen spezifischen Mechanismus entwickelt. Die Infektion neuer Zellen erfolgt über einen dünnen Infektionsschlauch, der, verglichen mit der Zelllänge, ungewöhnliche Länge erreichen kann (bis über 100 Mikrometer). Der Infektionsschlauch kann, wie eine Injektionsspritze, in Gewebe und Zellen eindringen, wo der Sporeninhalt durch einen massiven Druckanstieg innerhalb der Spore in den neuen Wirt überführt wird. Es werden normalerweise immer zwei Sporenarten gebildet, eine, die sehr schnell neue Zellen infiziert, und eine weitere, die dies erst zeitverzögert tut, bei einigen Mikrosporidienarten sind diese auch morphologisch unterscheidbar.
Die kompakt gebaute, meist runde, ovale, birnen- oder stäbchenförmige Spore besitzt eine dicke Zellwand, die zweilagig aufgebaut ist. Die innere (endospore) Lage besteht aus Proteinen und Chitin, die äußere nur aus Proteinen. Dabei dient die innere Lage vermutlich vor allem dazu, den extremen Druck, der bei der Infektion aufgebaut wird, zu widerstehen (es werden mehr als 7 Megapascal erreicht), die äußere dient zum Zellkontakt, zum Beispiel zur Wirtserkennung. Dazu besitzen sie oft stachelartige Fortsätze. Im Inneren der Spore befindet sich, neben dem Zellkern und wenig Cytoplasma, vor allem der Injektionsapparat, der den Infektionsschlauch ausbildet, und eine Polaroplast genannte Speicherorganelle für Membranbestandteile. Der Überdruck bei der Infektion wird durch eine Vakuole im hinteren Abschnitt erzeugt, die dazu rasch an Größe zunimmt und so Druck aufbaut. Der Infektionsschlauch am vorderen Ende der Spore wirkt fadenförmig, daher auch Polarfilament genannt, er ist meist schleifenartig aufgerollt. Beim Bienenparasiten Nosema apis ist die Spore 5 Mikrometer lang, der Injektionsschlauch (Polarfilament) in ihrem Inneren erreicht 300 Mikrometer Länge. Etwa ein Fünftel der Mikrosporidien-Gattungen besitzen stattdessen einen kurzen, stabartigen Injektionsschlauch, der nur etwa Sporenlänge erreicht. Bei ihnen ist er in einer pilzförmigen Struktur, der Polkappe, verankert. Hauptbestandteil des Infektionsschlauchs ist ein PTP1 genanntes Protein. Dieses erweist sich bei Wirten (auch beim Menschen) als starkes Antigen, so dass die gegen es gerichtete Immunantwort den Wirt resistent gegen Infektionen machen kann. Beim Infektionsvorgang wird der Zellinhalt durch die expandierende Vakuole in der Spore in den Schlauch gedrückt. Der Polaroplast, der im Ruhezustand vor allem aus eingefalteter Membran besteht, stellt das notwendige Material für die neue Zellmembran im Schlauch zur Verfügung. Der Infektionschlauch durchdringt angrenzendes Gewebe und andere Hindernisse, unter Einschluss möglicher anderer Mikrosporidien-Sporen, die im Weg wären.
Zahlreiche Arten von Mikrosporidien erzeugen mehrere Sporenformen, meist zwei (dimorph), selten drei oder mehr (polymorph). Unterschiedliche Formen dienen entweder alternativ der schnellen Infektion innerhalb eines Wirts und der verzögerten Keimung zur Infektion neuer Wirte, oder es werden bei verschiedenen Wirtsarten unterschiedliche Sporenformen ausgebildet. Bei anderen sehen alle Sporen gleich aus und unterscheiden sich nur in der Schnelligkeit der Keimbereitschaft. Neue Wirte werden durch langsam keimende Sporen infiziert, meist nach Aufnahme über den Mund und Verdauungstrakt (oral). Bei wasserlebenden Mikrosporidien gibt es hyperparasitische Arten, bei denen der Infektionsschlauch die Körper- oder Darmwand des Wirts durchdringt, um den im Inneren lebenden Parasiten zu parasitieren.
Die neu gebildeten Sporen gelangen meist durch den Kot, den Urin, oder erst nach dem Tod des Wirtes ins Freie. Bei vielen Wirten, darunter den meisten Wirbeltieren, verläuft die Infektion chronisch, so dass über lange Zeiträume Sporen ausgeschieden werden. Bei Insekten kommt es, nach verzögertem und oft relativ symptomlosen Beginn, auch zu schwerer verlaufenden Infektionen, die mit dem Tod des Wirts enden können. Auch bei den chronisch verlaufenden Infektionen ist meist die Lebensdauer und der Allgemeinzustand der Wirtsarten beeinträchtigt.
Die meisten Mikrosporidien-Arten sind wirtsspezifisch, sie befallen nur eine einzige Wirtsart oder wenige nahe verwandte Arten. Innerhalb des Wirts sind sie in der Regel auf bestimmte Gewebe spezialisiert. Mikrosporidien der Gattung Nosema, die Erreger der Nosemose, sind nicht nur spezialisiert auf Honigbienen der Gattung Apis, sondern befallen in diesen ausschließlich Zellen des Mitteldarms. Oft besitzen nahe verwandte Wirtsarten ihre jeweils eigenen, ebenso nahe verwandten Parasiten (genannt „Ko-Kladogenese“). In vielen Fällen ist die Wirtsspezifität durch Umweltfaktoren vorgegeben, d. h. unter experimentellen Bedingungen, im Labor, können auch Arten infiziert werden, bei denen dies im Freiland nie auftritt. Andere Arten besitzen ein weites Wirtsspektrum, z. B. alle Säugetiere, die sie opportunistisch nutzen.
Mikrosporidien-Infektionen gehören zu den häufigsten parasitischen Erkrankungen im Tierreich. Bei etwa der Hälfte der Stämme des Tierreichs wurden Mikrosporidien als Parasiten registriert. Besser untersucht sind aber nur die, relativ wenigen, Tiergruppen, bei denen die Infektion zu wirtschaftlichen oder gesundheitlichen Schäden führen, die für den Menschen bedeutsam sind. Es wird daher angenommen, dass weitaus die meisten Arten bis heute unentdeckt und unbeschrieben sind. Die meisten bekannten Wirtsarten gehören zu den Insekten und den Krebstieren. Mikrosporidien können erhebliche Schäden anrichten: Die Pébrine-Krankheit durch Nosema bombycis führte etwa Mitte des 19. Jahrhunderts zum Zusammenbruch der europäischen Seidenraupen-Zucht. Andere Arten, etwa Parasiten von Stechmücken besitzen aber als Regulatoren oder im Rahmen der biologischen Schädlingsbekämpfung auch positive Wirkungen für den Menschen. Etwa 160 Arten aus 17 Gattungen infizieren Fischarten. Beim Menschen als Wirt wurden 14 Mikrosporidien registriert, davon keine einzige wirtsspezifische. Arten der Gattung Endoreticulatus sind etwa opportunistische Erreger, die sowohl bei Menschen und anderen Wirbeltieren als auch bei Insekten-Arten vorkommen.
Microsporidiose ist der Oberbegriff für Krankheitsbilder durch diese Organismen.
Der nomenklatorische Status der Microsporidia ist nicht eindeutig. Sie wurden als Stamm nach den Internationalen Regeln für die Zoologische Nomenklatur (ICZN) behandelt (damals den Sporozoa zugeordnet), es herrscht jedoch Unklarheit über die Autor-Zitierung. Auch ist fraglich, ob der Name nach dem Internationalen Code der Botanischen Nomenklatur (ICN) gültig ist. Die Zugehörigkeit der Microsporidia zu den Pilzen und daher auch die Zuständigkeit des ICN hat sich 2007 ergeben. Die Zuordnung zu Balbiani (C. R. Acad. Sci Paris 95: S. 1168, 1882) ist daher vorläufig[1], aber bis heute üblich.
Die Abteilung (oder Stamm, Phylum) Microsporidia bilden mit einer Klade von wenig bekannten Organismen um die Gattung Rozella (als Rozellida oder Cryptomycota bezeichnet) und den Aphelidea, einer artenarmen Gruppe von Parasiten einzelliger Algen, eine Gruppe, die Opisthosporidia benannt worden ist[2][3], sie wird im klassischen System als Überstamm (Superphylum) eingeordnet. Die Opisthosporidia gehören zur Gruppe der Pilze (im weiteren Sinne).
Inzwischen wurde eine Reihe von Protisten gefunden, die zahlreiche Merkmale mit den Microsporidia gemeinsam hat, aber in einigen Merkmalen von diesen abweicht. Unter anderem existieren darin Vertreter mit weniger stark reduziertem Genom, die auch funktionsfähige Mitochondrien aufweisen. Zu dieser Gruppe zählen die Gattungen Mitosporidium, Paramicrosporidium und Nucleophaga. Eine weitere Gruppe, unter anderem Amphiamblys und Amphiacantha, als „Metchnikovelliden“ zusammengefasst, sind Hyperparasiten in, selbst parasitischen, Einzellern der Apicomplexa. Genetisch ähnlich sind zudem zahlreiche, bisher noch unbekannte Organismen, von denen nur ihre aus Umwelt-DNA gewonnene Basensequenz bekannt ist, ohne dass der zugehörige Organismus und dessen Biologie bisher bekannt geworden wäre; diese sind in Umweltproben teilweise sehr häufig. Fast die gesamte Gruppe, oft als Rozellomycota oder Cryptomycota zusammengefasst, erwies sich bei neueren Analysen als näher mit den Microsporidia als mit Rozella verwandt, so dass eine erweiterte Gruppe der Microsporidia unter Einschluss dieser Gruppen vorgeschlagen worden ist.[4]
Die Gliederung der Microsporidia in Gruppen ist nach bisherigem Wissensstand ungeklärt. Ein Vorschlag, sie, auch gestützt auf genetische Daten, in drei Klassen der Aquasporidia (vor allem im Süßwasser), Marinosporidia (vor allem marin) and Terresporidia (vor allem in terrestrischen Habitaten) einzuteilen[5] hat, aufgrund abweichender neuere Daten dazu, letztlich keine Akzeptanz gefunden.
In den 1850er Jahren verwüstete die Pébrine-Krankheit die europäische Seidenraupenzucht. Der Schweizer Forscher Carl Wilhelm von Nägeli entdeckte im Jahr 1857 darin infektiöse „Globuli“, die er als Nosema bombycis beschrieb. Ihre Lebensweise wurde dann 1870, vor allem durch den berühmten Louis Pasteur aufgeklärt und Gegenmaßnahmen empfohlen, so dass sich die Industrie erholte. Nägeli ordnete seinen Fund in die „Spaltpilze“ oder Schizomycetes, eine heute aufgegebene Zusammenfügung untereinander nicht näher verwandter „niederer“ Pilze und Bakterien, ein. Édouard-Gérard Balbiani stellte 1882 Nosema und Verwandte in die Gruppe der Sporozoa und prägte die Bezeichnung Microsporidia („microsporidies“). Er stellte für sie eine Gruppe der „Cnidosporidia“ auf, die außerdem einige nach heutiger Kenntnis nicht verwandte Gruppen, unter anderem die Myxozoa (nach heutiger Kenntnis Nesseltiere) umfasste.
Weitere Erkenntnisse zu den Microsporidia gelangen dann erst nach Erfindung der Elektronenmikroskopie in den 1950er Jahren. Nun wurde klar, dass den Microsporidia zahlreiche Merkmale fehlen, die sonst innerhalb der Tiere fast universell verbreitet sind. So kommen in den Zellen weder Mitochondrien noch ein Golgi-Apparat oder Peroxisomen vor, in keinem Lebensstadium existieren begeißelte Zellen mit Flagellum oder davon ableitbare Strukturen. Thomas Cavalier-Smith entwickelt daraufhin die Hypothese, die Microsporidia gehörten zu einer (paraphyletischen) Gruppe urtümlicher Eukaryoten, die sich vom gemeinsamen Stammbaum abgespalten hätten, noch bevor diese Strukturen evolviert seien. Diese „Archezoa“ genannte Gruppierung wurde auch von den ersten genetischen Analysen gut unterstützt. Mitte der 1990er Jahre entdeckten dann zahlreiche Arbeitsgruppen dicht nacheinander Merkmale vor allem verschiedener Proteinfamilien, die stattdessen eine nähere Verwandtschaft der Microsporidia mit den Pilzen nahelegten. Letztlich erwies sich die Unterstützung für die Archezoa durch bessere genetische Methoden als sogenanntes „Long-branch attraction“-Artefakt, bei dem sehr stark von den übrigen Gruppen abweichende DNA-Sequenzen vom Sortieralgorithmus irrtümlich nach außen abgedrängt werden und so eine basale Stellung vortäuschen. Eine Zeitlang war nun unklar, ob die Microsporidia als Schwestergruppe der Pilze aufgefasst werden sollten, oder ob sie zu den Pilzen zugehörig seien. Die neue Position wurde erst nach 2010, durch weiter verbesserte Analysemethoden und neu entdeckte Organismen der Verwandtschaftsgruppe, geklärt.[6]
Die Microsporidia, auch Microspora oder Microsporea, sind einzellige, zu den Pilzen gehörende Parasiten und Erreger der Mikrosporidiasis. Sie können eine Größe von wenigen µm erreichen (2–12 µm). Microspora parasitieren meist intrazellulär bei Vertretern vieler Tierstämme, seltener in anderen Protisten aus der Sar genannten Klade der Eukaryoten. Eine Übertragung erfolgt durch Aufnahme einzelliger Sporen (meist oral, d. h. durch den Mund).
Microsporidia je grupa sporulirajućih jednoćelijskih parazita. Smatrane su protozoama ili protistima, ali se sada zna da su gljive[5] ili sestrinska grupa gljiva.[6] Slobodno procjenjujući, ima ih 1500 vrsta, a vjerovatno više od milion[7] do sada imenovanih.
Microsporidia su ograničene na životinje domaćine, a sve glavne grupe životinja su om domaćini. Većina zarazi insekte, ali su odgovorni i za uobičajene bolesti rakova i riba. Navedene vrste microsporidia obično zaraze jednu vrstu domaćina ili grupu blisko srodnih taksona. Nekoliko vrsta, od kojih je većina oportunističkih, inficira i ljude.
Približno 10 % vrsta čine paraziti kičmenjaka – uključujući i ljude, kod kojih mogu izazvati mikrosporidijozu. Nakon infekcije, oni na različite načine utiču na svoje domaćine i napadaju sve organe i tkiva, mada općenito specifično različite vrste mikrosporidija. Neke su vrste smrtonosne, a neke se koriste u biološkoj kontroli insekata štetočina. Parazitskaa kastracija, gigantizam ili promjena spola domaćina, potencijalni su efekti mikrosporidijskog parazitizma (kod insekata). U najsavremenijim slučajevima parazitizma mikrosporidije u potpunosti ovladaju ćelijom domaćina i kontrolirau njen metabolizam i razmnožavanje, formirajući ksenome.[8] Replikacija se odvija unutar ćelija domaćina, koji su zaraženi jednoćelijskim sporama, koje su prečnika u rasponu od 1–40 µm, što ih čini jednim od najmanjih eukariota. Microsporidije koje inficiraju sisare imaju promjer oko 1,0–4,0 μm.[9] They also have the smallest eukaryotic genomes. Ermin "microsporidium" također se koristi kao zamjena za članove incertae sedis.[10]
Mikrosporidije nemaju mitohondrija, umjesto kojih imaju mitosome. Također im nedostaju strukture za kretanje, poput bičeva. Mikrosporidije stvaraju visoko otporne spore, sposobne da prežive izvan svog domaćina i do nekoliko godina. Morfologija spore korisna je u razlikovanju različitih vrsta. Spore većine vrsta su ovalne ili kruškolike, ali i šipkaste ili sferne spore nisu neuobičajene. Nekoliko rodova ima spore jedinstvenog oblika za taj rod.
Spora je zaštićena zidom koji se sastoji od tri sloja:
U većini slučajeva postoje dva usko povezana jedra, tvoreći "diplokarion", ali ponekad postoji samo jedan.
Prednja polovina spore sadrži aparat sličan harpunu s dugačkim navojem „polarnog vlakna“ koji je namotan u stražnjoj polovini spora. Prednji dio polarne niti je okružen "polaroplastom", lamelnom membranom. Iza polarnog vlakna nalazi se stražnja vakuola.[8]
U crijevima domaćina spora klija, stvarajući osmotski pritisak sve dok mu kruti dio zida ne ošteti na najtanjem mjestu na vrhu. Stražnja vakuola nabubri, prisiljavajući polarni filament da brzo izbaci infektivni sadržaj u citoplazmu potencijalnog domaćina. Istovremeno se materijal vlakna preuređuje tako da formira cijev koja djeluje kao hipodermna igla i prodire u epitel crijeva.
Unutar ćelije domaćina, raste sporoplazma, dijeli ili formira multinukleat plazmodija, prije stvaranja novih spora. Životni ciklus se među njima znatno razlikuje. Neki imaju jednostavan aseksualni životni ciklus,[12] dok drugi imaju složen, koji uključuje više domaćina i aseksualnih i seksualnih reprodukcija. Različite vrste spora mogu se stvarati u različitim fazama, vjerojatno s različitim funkcijama, uključujući autoinfekciju (prijenos unutar jednog domaćina).
U životinja i ljudi mikrosporidije često uzrokuju hronične, oslabljujuće bolesti a ne smrtonosne infekcije. Učinci na domaćina uključuju smanjenu dugovječnost, plodnost, težinu i opću energičnost. Često se izvještava o vertikalnom prijenosu mikrosporidija. U slučaju insekata domaćina, vertikalni prijenos često se događa transovarijski, tako da mikrosporidijski paraziti prelaze iz jajnika ženke domaćina u jajašce i na kraju se razmnožavaju u zaraženim larvama. Amblyospora salinaria n. sp. koji zarazi komarca Culex salinarius Coquillett i Amblyospora californica koji zarazi komarcem Culex tarsalis Coquillett, navode se kao tipski primjeri transovarijskog prijenosa mikrosporidija.[13][14][15][16]
Mikrosporidije, tačnije infekcije komaraca Vavraia culicis, istražuju se kao mogući obrazac razvoja mjera protiv malarij.[17] Mikrosporidijska infekcija Anopheles gambiae (glavni vektor malarije je Plasmodium falciparum) smanjuje malarijsku infekciju unutar komaraca i skraćuje im životni vijek.[18]
Kako većina komaraca zaraženih malarijom prirodno umire prije nego što je parazit malarije dovoljno zreo za prijenos, svako povećanje smrtnosti komaraca putem mikrosporidijanske infekcije može umanjiti prijenos malarije na ljude.
Mikrosporidijske infekcije ljudi ponekad uzrokuju bolest koja se zove mikrosporidioza. Najmanje 14 mikrosporidijanskih vrsta, iz osam rodova, prepoznate su kao ljudski patogeni. Oni uključuju Trachipleistophora hominis.[19]
Mikrosporidije mogu zaraziti različite domaćine, uključujući i one koji su i sami paraziti. U tom slučaju, mikrosporidijska vrsta jeste hiperparazit, tj. parazit parazita. Kao primjer, poznato je više od osamnaest vrsta koje parazitiraju a Digenea (parazitske pljosnate gliste). Ove gliste su i same paraziti u raznim kičmenjacima i mehkušci ma. Osam ovih vrsta pripada rodu Nosema .[20]
Mikrosporidije imaju najmanje poznate (nuklearne) eukariotske genom e. Parazitski način života mikrosporidija doveo je do gubitka mnogih mitohondrija l i Golgijevih gena, pa čak su i njihove ribosomnne RNK manje u usporedbi s onima kod većine eukariota. Kao posljedica toga, genomi mikrosporidija su mnogo manji nego kod ostalih eukariota. ]]Do sada poznati mikrosporidijski genomi su veličine od 2,5 do 11,6 [[bazni par|megabaza, a kodiraju od 1.848 do 3.266 proteina što je u istom rasponu koliko i kod bakterija.[21] Izgleda da se u mikrosporidiji događalo više puta. Na primjer, genomi Encephalitozoon romaleae i Trachipleistophora hominis sadrže gene koji potiču od životinja i bakterija, a neki čak i iz gljiva. Izgleda da se kod mikrosporidija više puta dogodio horizontalni transfer gena (HTG). Naprimjer, genomi Encephalitozoon romaleae i Trachipleistophora hominis sadrže derivirane gene životinja i bakterija, a kod nekih, čak i gljiva.[21]
Prvi opisani rod mikrosporidija, Nosema, inicijalno je Nägeli stavio u skupinu gljiva Schizomycetes zajedno s nekim bakterijama i kvascima.[23][24] Neko vrijeme mikrosporidije su smatrane vrlo primitivnim eukariotima, smještenim u protozojsku skupinu Cnidospora.[3] Kasnije, posebno zbog nedostatka mitohondrija, premještene su zajedno s ostalim Protozoaama poput Diplomonada, Parabasalida i Archamoebae u protozojsku grupu Archezoa.[25] Novija istraživanja odbacila su ovu teorij ranog porijekla (za sve navedeno). Ipak, predlažu se da su mikrosporidije visoko razvijeni i specijalizovani organizmi, koji jednostavno više nemaju potrebne funkcije, jer ih opskrbljuje domaćin.[26] Nadalje, organizmi koji stvaraju spore uopće imaju složen sistem razmnožavanja, i seksualni i aseksualni, koji izgledaju daleko od primitivnih.
Danas su mikrosporidije smještene unutar gljiva ili kao sestrinska grupa gljiva sa zajedničkim predakom.[27][28][29][30]
Formiranje kladusa uglavnom se temelji na staništu i domaćinu. Vossbrinck i Debrunner-Vossbrinck, na osnovu staništa, predlažu tri klase mikrosporidija: Aquasporidia, Marinosporidia i Terresporidia.[31] Druga klasifikacija, mogla bi biti:[32]
Treća reklasifikacija jeCavalier-Smith 1993:[33]
|journal= (pomoć) |journal= (pomoć) Microsporidia je grupa sporulirajućih jednoćelijskih parazita. Smatrane su protozoama ili protistima, ali se sada zna da su gljive ili sestrinska grupa gljiva. Slobodno procjenjujući, ima ih 1500 vrsta, a vjerovatno više od milion do sada imenovanih.
Microsporidia su ograničene na životinje domaćine, a sve glavne grupe životinja su om domaćini. Većina zarazi insekte, ali su odgovorni i za uobičajene bolesti rakova i riba. Navedene vrste microsporidia obično zaraze jednu vrstu domaćina ili grupu blisko srodnih taksona. Nekoliko vrsta, od kojih je većina oportunističkih, inficira i ljude.
Približno 10 % vrsta čine paraziti kičmenjaka – uključujući i ljude, kod kojih mogu izazvati mikrosporidijozu. Nakon infekcije, oni na različite načine utiču na svoje domaćine i napadaju sve organe i tkiva, mada općenito specifično različite vrste mikrosporidija. Neke su vrste smrtonosne, a neke se koriste u biološkoj kontroli insekata štetočina. Parazitskaa kastracija, gigantizam ili promjena spola domaćina, potencijalni su efekti mikrosporidijskog parazitizma (kod insekata). U najsavremenijim slučajevima parazitizma mikrosporidije u potpunosti ovladaju ćelijom domaćina i kontrolirau njen metabolizam i razmnožavanje, formirajući ksenome. Replikacija se odvija unutar ćelija domaćina, koji su zaraženi jednoćelijskim sporama, koje su prečnika u rasponu od 1–40 µm, što ih čini jednim od najmanjih eukariota. Microsporidije koje inficiraju sisare imaju promjer oko 1,0–4,0 μm. They also have the smallest eukaryotic genomes. Ermin "microsporidium" također se koristi kao zamjena za članove incertae sedis.
Microsporidia are a group of spore-forming unicellular parasites. These spores contain an extrusion apparatus that has a coiled polar tube ending in an anchoring disc at the apical part of the spore.[7] They were once considered protozoans or protists, but are now known to be fungi,[8] or a sister group to fungi.[9] These fungal microbes are obligate eukaryotic parasites that use a unique mechanism to infect host cells.[10] They have recently been discovered in a 2017 Cornell study to infect Coleoptera on a large scale. So far, about 1500 of the probably more than one million[11] species are named. Microsporidia are restricted to animal hosts, and all major groups of animals host microsporidia. Most infect insects, but they are also responsible for common diseases of crustaceans and fish. The named species of microsporidia usually infect one host species or a group of closely related taxa. Approximately 10 percent of the species are parasites of vertebrates —several species, most of which are opportunistic, can infect humans, in whom they can cause microsporidiosis.
After infection they influence their hosts in various ways and all organs and tissues are invaded, though generally by different species of specialised microsporidia. Some species are lethal, and a few are used in biological control of insect pests. Parasitic castration, gigantism, or change of host sex are all potential effects of microsporidian parasitism (in insects). In the most advanced cases of parasitism the microsporidium rules the host cell completely and controls its metabolism and reproduction, forming a xenoma.[12]
Replication takes place within the host's cells, which are infected by means of unicellular spores. These vary from 1–40 μm, making them some of the smallest eukaryotes. Microsporidia that infect mammals are 1.0–4.0 μm.[13] They also have the smallest eukaryotic genomes.
The terms "microsporidium" (pl. "microsporidia") and "microsporidian" are used as vernacular names for members of the group. The name Microsporidium Balbiani, 1884[14] is also used as a catchall genus for incertae sedis members.[15]
Microsporidia lack mitochondria, instead possessing mitosomes. They also lack motile structures, such as flagella.
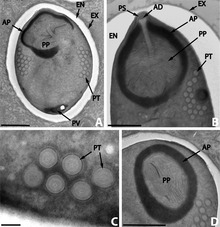
Microsporidia produce highly resistant spores, capable of surviving outside their host for up to several years. Spore morphology is useful in distinguishing between different species. Spores of most species are oval or pyriform, but rod-shaped or spherical spores are not unusual. A few genera produce spores of unique shape for the genus.
The spore is protected by a wall, consisting of three layers:
In most cases there are two closely associated nuclei, forming a diplokaryon, but sometimes there is only one.
The anterior half of the spore contains a harpoon-like apparatus with a long, thread-like polar filament, which is coiled up in the posterior half of the spore. The anterior part of the polar filament is surrounded by a polaroplast, a lamella of membranes. Behind the polar filament, there is a posterior vacuole.[12]
In the gut of the host the spore germinates; it builds up osmotic pressure until its rigid wall ruptures at its thinnest point at the apex. The posterior vacuole swells, forcing the polar filament to rapidly eject the infectious content into the cytoplasm of the potential host. Simultaneously the material of the filament is rearranged to form a tube which functions as a hypodermic needle and penetrates the gut epithelium.
Once inside the host cell, a sporoplasm grows, dividing or forming a multinucleate plasmodium, before producing new spores. The life cycle varies considerably. Some have a simple asexual life cycle,[17] while others have a complex life cycle involving multiple hosts and both asexual and sexual reproduction. Different types of spores may be produced at different stages, probably with different functions including autoinfection (transmission within a single host).
In animals and humans, microsporidia often cause chronic, debilitating diseases rather than lethal infections. Effects on the host include reduced longevity, fertility, weight, and general vigor. Vertical transmission of microsporidia is frequently reported.
In the case of insect hosts, vertical transmission often occurs as transovarial transmission, where the microsporidian parasites pass from the ovaries of the female host into eggs and eventually multiply in the infected larvae. Amblyospora salinaria n. sp. which infects the mosquito Culex salinarius Coquillett, and Amblyospora californica which infects the mosquito Culex tarsalis Coquillett, provide typical examples of transovarial transmission of microsporidia.[18][19][20][21] Microsporidia, specifically the mosquito-infecting Vavraia culicis, are being explored as a possible 'evolution-proof' malaria-control method.[22] Microsporidian infection of Anopheles gambiae (the principal vector of Plasmodium falciparum malaria) reduces malarial infection within the mosquito, and shortens the mosquito lifespan.[23] As the majority of malaria-infected mosquitoes naturally die before the malaria parasite is mature enough to transmit, any increase in mosquito mortality through microsporidian-infection may reduce malaria transmission to humans. In May 2020, researchers reported that Microsporidia MB, a symbiont in the midgut and ovaries of An. arabiensis, significantly impaired transmission of P. falciparum, had "no overt effect" on the fitness of host mosquitoes, and was transmitted vertically (through inheritance).[24]
Microsporidian infections of humans sometimes cause a disease called microsporidiosis. At least 14 microsporidian species, spread across eight genera, have been recognized as human pathogens. These include Trachipleistophora hominis.[25]
Microsporidia can infect a variety of hosts, including hosts which are themselves parasites. In that case, the microsporidian species is a hyperparasite, i.e. a parasite of a parasite. As an example, more than eighteen species are known which parasitize digeneans (parasitic flatworms). These digeneans are themselves parasites in various vertebrates and molluscs. Eight of these species belong to the genus Nosema.[26] Similarly, the microsporidian species Toguebayea baccigeri is a parasite of a digenean, the faustulid Bacciger israelensis, itself an intestinal parasite of a marine fish, the bogue Boops boops (Teleostei, Sparidae).[27]
Microsporidia have the smallest known (nuclear) eukaryotic genomes. The parasitic lifestyle of microsporidia has led to a loss of many mitochondrial and Golgi genes, and even their ribosomal RNAs are reduced in size compared with those of most eukaryotes. As a consequence, the genomes of microsporidia are much smaller than those of other eukaryotes. Currently known microsporidial genomes are 2.5 to 11.6 Mb in size, encoding from 1,848 to 3,266 proteins which is in the same range as many bacteria.[28]
Horizontal gene transfer (HGT) seems to have occurred many times in microsporidia. For instance, the genomes of Encephalitozoon romaleae and Trachipleistophora hominis contain genes that derive from animals and bacteria, and some even from fungi.[28]
The first described microsporidian genus, Nosema, was initially put by Nägeli in the fungal group Schizomycetes together with some bacteria and yeasts.[29][30] For some time microsporidia were considered as very primitive eukaryotes, placed in the protozoan group Cnidospora.[1] Later, especially because of the lack of mitochondria, they were placed along with the other Protozoa such as diplomonads, parabasalids and archamoebae in the protozoan-group Archezoa.[31] More recent research has falsified this theory of early origin (for all of these). Instead, microsporidia are proposed to be highly developed and specialized organisms, which just dispensed functions that are needed no longer, because they are supplied by the host.[32] Furthermore, spore-forming organisms in general do have a complex system of reproduction, both sexual and asexual, which look far from primitive.
Since the mid-2000s microsporidia are placed within the Fungi or as a sister-group of the Fungi with a common ancestor.[33][34][35][36]
Work to identify clades is largely based on habitat and host. Three classes of Microsporidia are proposed by Vossbrinck and Debrunner-Vossbrinck, based on the habitat: Aquasporidia, Marinosporidia and Terresporidia.[37]
A second classification by Cavalier-Smith 1993:[38]
Microsporidia are a group of spore-forming unicellular parasites. These spores contain an extrusion apparatus that has a coiled polar tube ending in an anchoring disc at the apical part of the spore. They were once considered protozoans or protists, but are now known to be fungi, or a sister group to fungi. These fungal microbes are obligate eukaryotic parasites that use a unique mechanism to infect host cells. They have recently been discovered in a 2017 Cornell study to infect Coleoptera on a large scale. So far, about 1500 of the probably more than one million species are named. Microsporidia are restricted to animal hosts, and all major groups of animals host microsporidia. Most infect insects, but they are also responsible for common diseases of crustaceans and fish. The named species of microsporidia usually infect one host species or a group of closely related taxa. Approximately 10 percent of the species are parasites of vertebrates —several species, most of which are opportunistic, can infect humans, in whom they can cause microsporidiosis.
After infection they influence their hosts in various ways and all organs and tissues are invaded, though generally by different species of specialised microsporidia. Some species are lethal, and a few are used in biological control of insect pests. Parasitic castration, gigantism, or change of host sex are all potential effects of microsporidian parasitism (in insects). In the most advanced cases of parasitism the microsporidium rules the host cell completely and controls its metabolism and reproduction, forming a xenoma.
Replication takes place within the host's cells, which are infected by means of unicellular spores. These vary from 1–40 μm, making them some of the smallest eukaryotes. Microsporidia that infect mammals are 1.0–4.0 μm. They also have the smallest eukaryotic genomes.
The terms "microsporidium" (pl. "microsporidia") and "microsporidian" are used as vernacular names for members of the group. The name Microsporidium Balbiani, 1884 is also used as a catchall genus for incertae sedis members.
 Xenoma on flatfish caused by Glugea stephani
Xenoma on flatfish caused by Glugea stephani
Los microsporidios (Microsporidia o también Microsporidiomycetes)[1] son un conjunto de hongos que son parásitos intracelulares de animales y algunos protozoos.[2] Se han descrito unas 800 especies.
Fueron clasificados como protozoos, como protistas y como hongos. En realidad son un grupo basal de hongos parásitos obligados que descienden de organismos unicelulares similares a las rozelas, que en el proceso evolutivo de su adaptación a la vida parasitaria, han perdido las mitocondrias.
Estudios filogenéticos recientes proponen clasificar a Rozellomycetes junto con Microsporidia en una división Rozellomycota, ya que según los estudios Rozellomycetes podría ser un grupo parafilético de los microsporidios. Varios análisis moleculares confirman que Microsporidia y Rozellomycetes comprenden grupos hermanos, mientras que el otro grupo Aphelidiomycota se había colocado con ellos en el taxón Opisthosporidia, pero estudios recientes han encontrado que está más emparentados con Eumycota.[3][4][5]
Los microsporidios, como su nombre indica, se caracterizan por sus esporas de tamaño mínimo, de 2 a 20 µm, que contienen el esporoplasma (el parásito infectivo) y un filamento polar sencillo. Una de las paredes de la espora contiene quitina, otro rasgo de los hongos. Presentan paredes celulares quitinosas como los hongos, aunque esta puede perderse durante la absorción, debido al parasitismo.
Los parásitos maduros, trofozoides, en ocasiones, producen zigotos por meiosis, que posteriormente pueden producir más esporas. La reproducción puede ser sexual o asexual. Presentan zoosporas uniflageladas opistocontas que tienen un rizoplasto sólido asociado con un cinetosoma largo. Penetran las células huésped a través de un tubo germinal y absorben el contenido interno de los orgánulos del huésped obteniendo a través de ellos los nutrientes.
Los microsporidios son parásitos intracelulares obligados de diversas especies animales, que infectan tanto vertebrados como invertebrados y algunos protozoos. El término microsporidio es una designación común, que se utiliza para nombrar a un conjunto de microorganismos caracterizados por la producción de esporas pequeñas y por la carencia de mitocondrias.
La ausencia de plastos o mitocondrias hizo pensar que se trataba de formas muy primitivas descendientes separados de los primeros eucariontes, a los que se llamó arquezoos, anteriores a la simbiosis que dio origen a esos orgánulos. Llamó la atención que los ribosomas del microsporidio Vairimorpha necathrix mostraran dimensiones típicas de los de las bacterias, no de eucariontes. El análisis filogenético de los genes ribosómicos y de otros, ha demostrado que el grupo está formado por varias derivaciones independientes, reducidas por parasitismo, de formas basales del reino Fungi. Un examen más detallado del ARN ribosómico confirmó que se trataba de una reducción típica del parasitismo.
La microsporidiosis puede causar una variedad de enfermedades en el ser humano involucrando múltiples órganos incluyendo: intestino, ojos, pulmón, de senos paransales, riñón y músculos; tanto en inmunocomprometidos como en inmunocompetentes. Después del aparato gastrointestinal, la microsporidiosis ocular es la manifestación más frecuente en humanos. Las dos entidades clínicas oftalmológicas de la microsporidiosis son: la infección estromal corneal profunda en pacientes inmunocompetentes y la queratoconjuntivitis superficial en pacientes con SIDA2 y más recientemente también encontrada en pacientes inmunocompetentes Las manifestaciones oculares incluyen queratoconjuntivitis puntiforme superficial y queratitis estromal, que dependerá del estado inmunológico de paciente.
La queratoconjuntivitis usualmente se encuentra en pacientes inmunocomprometidos o en pacientes con lentes de contacto; principalmente por el gen Encephalitozoon, mientras que la queratitis estromal es causada por Nosema y Microsporidium. Sin embargo, reportes recientes han sugerido que la queratoconjuntivitis también puede ocurrir en pacientes inmunocompetentes. Davis et cols, reportaron un caso de queratitis estromal causada por la microsporidiosis en un paciente sano, sin antecedente de trauma. Pinnolis et cols y Ashton et cols describieron dos casos de involucro corneal en un niño de 11 años de edad de Sri Lanka con una úlcera corneal, y en una mujer de 26 años de edad de Botsuana con una úlcera corneal perforada, ambos pacientes, sin antecedentes de importancia y con serología negativa para VIH.
Los microsporidios parasitan las células intestinales de mamíferos, peces y aves, aunque también se han encontrado en invertebrados (Nosema produce la pebrina, una enfermedad de los gusanos de seda Bombyx mori, como describió Luis Pasteur). Los microsporidios aparte de los animales, también parasitan algunos protozoos como los ciliados.
Nosema apis es un microsporidio que causa una enfermedad denominada nosemosis, que ataca a la abeja melífera, causándole inflamación intestinal y diarrea. El antibiótico utilizado en el tratamiento de esta enfermedad es la fumagilina.
La taxonomía propuesta para los microsporidios es aproximadamente la siguiente. Aunque los microsporidios se consideren hongos deberán seguir empleando el Código de Nomenclatura Zoológica usado tradicionalmente en los protistas debido al gran patrimonio taxonómico por el que han pasado.[5]
Los microsporidios (Microsporidia o también Microsporidiomycetes) son un conjunto de hongos que son parásitos intracelulares de animales y algunos protozoos. Se han descrito unas 800 especies.
Fueron clasificados como protozoos, como protistas y como hongos. En realidad son un grupo basal de hongos parásitos obligados que descienden de organismos unicelulares similares a las rozelas, que en el proceso evolutivo de su adaptación a la vida parasitaria, han perdido las mitocondrias.
Estudios filogenéticos recientes proponen clasificar a Rozellomycetes junto con Microsporidia en una división Rozellomycota, ya que según los estudios Rozellomycetes podría ser un grupo parafilético de los microsporidios. Varios análisis moleculares confirman que Microsporidia y Rozellomycetes comprenden grupos hermanos, mientras que el otro grupo Aphelidiomycota se había colocado con ellos en el taxón Opisthosporidia, pero estudios recientes han encontrado que está más emparentados con Eumycota.
Väikeeosloomad[1] ehk pisieosloomad[2] (Microsporidia) seente hõimkond, kuhu kuuluvad üherakulised spoore moodustavad parasiidid. Varem käsitleti neid hõimkonnana protistide riigist.
Väikeeosloomad ehk pisieosloomad (Microsporidia) seente hõimkond, kuhu kuuluvad üherakulised spoore moodustavad parasiidid. Varem käsitleti neid hõimkonnana protistide riigist.
Microsporidiomycota
Les Microsporidies, Microsporidiomycota[1], anciennement Microsporidia, sont une division de champignons parasites intracellulaires obligatoires, appartenant au règne des Fungi.
Il semblerait qu'ils soient génétiquement très proches des Eumycètes dont ils dérivent peut-être par adaptation au parasitisme (Une hypothèse proposée par James & al. (2006) est que le groupe des microsporidia dériverait d'un ancêtre chytride endoparasite (proche de Rozella allomycis) appartenant à la première branche divergente de l'arbre phylogénétique des Fungi)[2]. La comparaison des régions de synténie avec d'autres champignons montre que les microsporidies seraient proches des Mucoromycotina. On en connait maintenant 150 genres et 1200 espèces. Cependant, une nouvelle estimation du nombre d’espèces montre qu’il se pourrait que le nombre d’espèce de microsporidies atteigne le nombre d’espèces animales.
Les microsporidies présentent une adaptation caractérisée par la réduction. En effet les microsporidies sont réduites à beaucoup de niveaux par rapport aux autres eucaryotes ; au niveau du génome en premier lieu mais aussi au niveau de leur métabolisme (elles n'ont pas de mitochondries)[2].
Le génome de Encephalitozoon cuniculi a été entièrement séquencé. Ce parasite possède le génome eucaryote le plus petit connu à ce jour.
La première description d’une microsporidie fut faite au milieu du XIXe siècle, quand la pébrine décimait les vers à soie. Cette maladie causée par Nosema bombycis fut identifiée par Karl Wilhelm von Nägeli en 1857[3]. Ce champignon semble avoir été signalé pour la première fois en 1849, avant l'identification de la pébrine, par Guérin-Méneville, qui croyait erronément que les vers à soie sur lesquels il l'observait étaient atteints de muscardine[4]. Après l'identification de la pébrine, Cornalia, Franz Leydig[5], Balbiani[6] et Pasteur[7] étudièrent son agent microbien (« corpuscules ») de façon plus approfondie. C’est ensuite Louis Pasteur qui eut l’idée de sélectionner les individus non infectés pour recréer de nouveaux élevages sains.
Les Microsporidies parasitent de très nombreux Eucaryotes comme d'autres protistes, des Invertébrés, des Vertébrés dont l'Homme. Elles infectent aussi certains protistes tels que des ciliés qui sont elles-mêmes des parasites. Ces parasites de parasites s’expliquent par le fait que ces 2 espèces infectaient précédemment le même hôte et qu’une des deux espèces s’est adaptée pour parasiter le voisin. Les espèces du genre Nosema sont très pathogènes pour les Insectes. Nosema bomycis attaque le ver à soie (la chenille du bombyx du murier), Nosema apis attaque l'abeille adulte. De nombreux microsporidies sont des pathogènes de poissons et de crustacés. Les espèces des genres Enterocytozoon et Encephalitozoon ont pour réservoir divers animaux et peuvent être pathogène opportuniste de l'Homme, notamment chez les immunodéprimés. Chez l'Homme, les microsporidies peuvent provoquer des troubles intestinaux ou oculaires et peuvent être détectées par un examen parasitologique des selles.
Bien que répandu chez tous les animaux, il semble que les microsporidies soient plus présentes chez les poissons et les arthropodes. Elles peuvent être utilisées comme agents biologiques de contrôle contre certains insectes mais peuvent aussi décimer certaines apicultures et aquacultures.
Il existe 13 espèces de microsporidies qui infectent l’homme, dont Encephalitozoon cuniculi. Beaucoup de maladies sont associées à ces parasites et la prévalence en Europe est de 8 %.
Les microsporidies présentent une forme de résistance et de dissémination : la spore. La spore est le seul stade reconnaissable des microsporidies. C’est à ce stade que les espèces peuvent être différenciées et c’est le seul à être viable hors de la cellule hôte. La taille et la forme de la spore sont très conservés chez une même espèce.
La spore est entourée par une membrane plasmique classique ainsi que par deux parois extracellulaires rigides : exospore et endospore. L’exospore est faite d’une matrice glycoprotéique et fibreuse dense. L’endospore est composée de chitine alpha et d’autres protéines. Son épaisseur est assez uniforme sauf à l’apex où cette paroi est plus fine. À l’intérieur de la membrane se trouve le sporoplasme (cytoplasme de la spore) qui constitue le matériel infectieux. Il contient un seul noyau, il est riche en ribosomes et est occupé majoritairement par les trois structures nécessaires à l’infection:
Le polaroplaste est une grande organisation de membrane occupant la partie antérieure de la spore. La portion antérieure du polaroplaste est hautement organisée sous forme de membranes empilées appelées polaroplaste lamellaire alors que la portion postérieure est moins organisée et est appelée polaroplaste vésiculaire. L’organite jouant le rôle le plus évident dans l’infection est le filament polaire (ou tube polaire). Dans le sporoplasme, il est composé de couche de glycoprotéines ; 0.1 à 0.2 µm de diamètre et 50 à 500 µm de long. Il est attaché à l’apex via une structure en parapluie appelée disque d’ancrage. Sur 1/3 de la spore ce filament est raide et hélicoïdal (le nombre de spires et leur angle sont conservés et permettent d’identifier certaines espèces). Ce filament se termine au niveau de la vacuole postérieure. Il semblerait qu’il y ait un contact physique entre ces 2 structures.
Le cycle se déroule en trois phases : la phase infectieuse, la phase de prolifération et la phase de différenciation.
La phase infectieuse commence par la germination des spores qui est un événement biologique assez surprenant. Cette germination commence par un environnement favorable ; ces conditions varient selon les espèces mais sont peu connues. Il peut s’agir d’une altération de pH, de déshydratation suivie d’une réhydratation, de la présence d’anions, de cations, d’UV etc. Quand la germination commence, le premier signe est un grossissement de la spore et plus particulièrement du polaroplaste et de la vacuole postérieure. Il en résulte une forte augmentation de la pression osmotique. Cette augmentation de pression peut s’expliquer par deux mécanismes :
(L’augmentation du niveau de tréhalose pourrait aussi jouer le rôle de métabolite anti-stress.) Cependant on a constaté dans certaines espèces de microsporidies qu’il n’y avait pas de modification du niveau de tréhalose dans la cellule suggérant d’autres mécanismes pour l’entrée d’eau dans la spore durant la germination. Il existe une dernière hypothèse faisant intervenir la concentration de calcium dans l’influx d’eau et rôle de la calmoduline : la présence d’ions calcium améliore l’extrusion du tube polaire. Le début de dégradation de la membrane pourrait entraîner un afflux d’ions calcium dans le sporoplasme et ces ions induiraient l’influx d’eau mais aussi activeraient des enzymes telles que les tréhalases. Quoi qu’il en soit, l’augmentation de la pression osmotique entraîne la rupture du disque d’ancrage et la projection du filament polaire par éversion (ou dévagination : l'intérieur du tube passe à l'extérieur, comme retourner le doigt d’un gant sur lui-même). Cette éversion commence à l’apex, où le filament polaire casse la paroi au niveau où elle est la plus fine. Le filament devient un tube et l’intérieur passe à l’extérieur. C’est un évènement très rapide durant moins de 2 secondes et dont la vitesse est difficile à estimer. Le tube polaire mesure alors entre 50 et 500 µm (100 fois la taille de la spore) et le bout du tube peut aller à une vitesse de 100 µm/s. Ce tube sert donc de projectile et si une cellule est à côté, il peut transpercer la membrane plasmique. Une fois que le tube est totalement sorti, la pression osmotique résiduelle force le sporoplasme à passer dans le tube, qu’il parcourt en seulement 15 à 500 ms (notez la très forte déformation du sporoplasme). Ainsi le sporoplasme émerge directement dans le cytoplasme de la cellule hôte, il n’y a donc plus de reconnaissance en tant que corps étranger. La spore vide est abandonnée alors que le sporoplasme émerge dans la cellule avec une nouvelle membrane issue de la membrane du polaroplaste.
La durée d’évolution du cycle est toujours brève, 24 à 48h, mais elle est liée à plusieurs facteurs extrinsèques : la température ambiante, qui influe sur le nombre de spores obtenues, ou les tissus de l’hôte.
Une fois dans la cellule-hôte, le sporoplasme déposé se divise par scission binaire (mérogonie), parfois à l’intérieur d’une vacuole parasitophore (comme dans le cas d’Encephalitozoon cuniculi), en formes prolifératives (mérontes) avec une simple membrane plasmique. Ces mérontes possèdent de petits ribosomes proches de ceux des procaryotes et des organites parsemées. Le parasite induit certains changements non délétères chez l’hôte. Souvent la cellule se réorganise autour du parasite et tous les organites sont autour de lui.
Il s’agit de la formation des spores. Durant cette phase il y a une augmentation du réticulum endoplasmique et du nombre de ribosomes qui se regroupent en polyribosome. Le développement du sporonte consiste en plusieurs divisions et un développement du système d’extrusion (filament polaire, polaroplaste, vacuole postérieure). L'appareil de golgi donnerait naissance au filament polaire et le réticulum endoplasmique aux membranes du polaroplaste. Quand la maturité approche, la paroi de chitine de l’endospore se développe. L’élargissement de la vacuole parasitophore et/ou de la cellule hôte par les parasites conduit à la rupture de la cellule-hôte et au relargage des spores dans l’espace extracellulaire. Il y a ensuite libération de la spore complète qui va ensuite soit infecter le même hôte ou alors être transportée vers un autre hôte (via le système digestif). Les moyens de dissémination sont les urines, les fèces, la décomposition…
Les microsporidies sont des organismes anaérobies dénués de mitochondries, permettant l'oxydation des composés organiques. À la place, ils possèdent un organite appelé mitosome.
La difficulté des études biochimiques est due à leur mode de vie intracellulaire obligatoire. Concernant le métabolisme de base des microsporidies, on sait maintenant que ces parasites, ne possédant pas de cycle de Krebs, importent l’ATP de l’hôte. Cependant, elles produisent des acides lactiques et pyruviques et possèdent des enzymes impliquées dans la glycolyse, la voie des pentoses-phosphates et la voie de biosynthèse du tréhalose (et aussi pour sa dégradation. Contrairement aux êtres sans mitochondrie, les microsporidies ont une PDH (pyruvate déshydrogénase) et non une PFOR (pyruvate ferredoxin oxydoréductase). En revanche, la PDH aurait un rôle particulier dans les microsporidies car elle fonctionne avec uniquement la sous-unité E1 : elle utilise directement l’intermédiaire HETPP pour transférer les électrons à la ferodoxine. Il y a une autre différence avec les autres êtres sans mitochondrie : la phosphofructokinase serait ATP-dépendante comme chez les champignons au lieu d’être pyrophosphate dépendante.
Le fait que les microsporidies possèdent une HSP70 et une PDH et les données du génome d’Encephalitozoon cuniculi démontrent bien qu’elles dérivent d’un champignon possédant une mitochondrie.
Les microsporidies peuvent infecter de nombreux hôtes, parmi lesquels certains sont eux-mêmes des parasites. Dans ces cas, la microsporidie est un hyperparasite, c'est-à-dire un parasite de parasite. On connaît par exemple 18 espèces qui parasitent les digènes (un groupe de Plathelminthes parasites). Ces digènes sont eux-mêmes parasites de vertébrés et de mollusques. Huit de ces espèces appartiennent au genre Nosema[10]. De la même manière, la microsporidie Toguebayea baccigeri est hyperparasite du trématode digène Bacciger israelensis (Faustulidae), lui-même un parasite intestinal du bogue, Boops boops (Teleostei, Sparidae)[11].
D’abord classé dans les schizomycètes (sorte de « fourre-tout » pour les bactéries, levures…) ces parasites ont vu la création d’un nouveau groupe, les microsporidies, par Édouard-Gérard Balbiani en 1882. La place des microsporidies dans l’évolution a souvent changé depuis leur description au milieu du XIXe siècle. Elles ont d’abord été classées comme des « yeast-like fungus » puis, considérant leur mode d’infection unique, elles ont été exclues du groupe des champignons sans pour autant être rattachées à un autre groupe d’eucaryotes. Au fil des observations et classifications, elles ont été classées comme parasite, levure etc. C’est en 1983 que s’est posé une nouvelle fois l’attention de biologistes sur ces microsporidies. On les a alors considérées comme des eucaryotes primitifs puisqu’elles ne possèdent pas de mitochondrie, elles auraient alors divergé avant l’endosymbiose à l’origine des mitochondries. De plus les premières observations de phylogénie moléculaires ont montré que les microsporidies possédaient un ARN ribosomique 5.8 S fusionné avec l'équivalent de l’ARN 28S donnant ainsi un 23S comme chez les procaryotes. De même, plusieurs délétions sont présentes dans les ARNr en comparaison avec les ARNr conservées d’autres eucaryotes et procaryotes. Cependant, cette hypothèse a été remise en cause lors de la découverte de séquences nucléotidiques microsporidiales homologues aux gènes codant des protéines mitochondriales et d’autre analyses comme la phylogénie des tubulines alpha et beta ont montré qu’elles appartenaient au même groupe que les champignons. Le séquençage du génome d'Encephalitozoon cuniculi a confirmé cette hypothèse.
Concernant l’absence de mitochondrie, la présence de gène mitochondriaux prouve que les microsporidies en possédaient une mais qu’elle a été perdue. De nos jours les microsporidies sont considérées comme des champignons évolués proches des Mucoromycotina et très spécialisés dans le parasitisme et que leur simplicité apparente est due à une évolution réductrice causée par leur mode de vie particulier.
Selon NCBI (24 juin 2013)[12] :
Microsporidiomycota
Les Microsporidies, Microsporidiomycota, anciennement Microsporidia, sont une division de champignons parasites intracellulaires obligatoires, appartenant au règne des Fungi.
Il semblerait qu'ils soient génétiquement très proches des Eumycètes dont ils dérivent peut-être par adaptation au parasitisme (Une hypothèse proposée par James & al. (2006) est que le groupe des microsporidia dériverait d'un ancêtre chytride endoparasite (proche de Rozella allomycis) appartenant à la première branche divergente de l'arbre phylogénétique des Fungi). La comparaison des régions de synténie avec d'autres champignons montre que les microsporidies seraient proches des Mucoromycotina. On en connait maintenant 150 genres et 1200 espèces. Cependant, une nouvelle estimation du nombre d’espèces montre qu’il se pourrait que le nombre d’espèce de microsporidies atteigne le nombre d’espèces animales.
Les microsporidies présentent une adaptation caractérisée par la réduction. En effet les microsporidies sont réduites à beaucoup de niveaux par rapport aux autres eucaryotes ; au niveau du génome en premier lieu mais aussi au niveau de leur métabolisme (elles n'ont pas de mitochondries).
Le génome de Encephalitozoon cuniculi a été entièrement séquencé. Ce parasite possède le génome eucaryote le plus petit connu à ce jour.
Microsporidia merupakan kelompok parasit pembentuk-spora uniseluler. Mereka pernah dianggap protozoa atau protista, tetapi sekarang dikenal sebagai fungi,[6] atau kelompok saudara fungi.[7] 1500 mungkin lebih dari satu juta[butuh rujukan] spesies diberi nama. Microsporidia terbatas pada inang hewan, dan semua kelompok utama binatang merupakan tuan rumah microsporidia. Kebanyakan menginfeksi serangga, tetapi ada yang termasuk golongan krustasea dan ikan. Spesies bernama microsporidia biasanya menginfeksi satu spesies. Beberapa spesies, yang sebagian besar adalah oportunistik, juga menginfeksi manusia.
Microsporidia merupakan kelompok parasit pembentuk-spora uniseluler. Mereka pernah dianggap protozoa atau protista, tetapi sekarang dikenal sebagai fungi, atau kelompok saudara fungi. 1500 mungkin lebih dari satu juta[butuh rujukan] spesies diberi nama. Microsporidia terbatas pada inang hewan, dan semua kelompok utama binatang merupakan tuan rumah microsporidia. Kebanyakan menginfeksi serangga, tetapi ada yang termasuk golongan krustasea dan ikan. Spesies bernama microsporidia biasanya menginfeksi satu spesies. Beberapa spesies, yang sebagian besar adalah oportunistik, juga menginfeksi manusia.
De Microsporidia of Microspora[1] vormen een stam van eencellige parasieten, nauw verwant aan schimmels. Er bestaan waarschijnlijk meer dan een miljoen soorten, waarvan er ongeveer 1500 zijn beschreven. Microsporidia leven als parasieten op dieren, voornamelijk insecten. Ongeveer een tiende van alle soorten infecteert gewervelden, waaronder mensen. Sommige soorten worden gebruikt om insectenplagen tegen te gaan.
Infectie vindt plaats door middel van sporen, die de cellen van de gastheer binnendringen. Deze sporen zijn maar 1 tot 40 micrometer groot en behoren daarmee tot de kleinste eukaryotische cellen. Microsporidia vormen daarnaast de groep met het kleinste genoom van alle eukaryoten.
Microsporidia hebben geen mitochondriën maar in plaats daarvan simpelere structuren, zogenaamde mitosomen. Ze hebben ook geen flagella of andere organellen die voor motiliteit kunnen zorgen. Veel soorten hebben niet een maar twee celkernen, dit wordt een diplokaryon genoemd.
De meeste soorten hebben peervormige of ovale sporen, maar ook langwerpige of ronde sporen komen voor. Aan de achterkant van een spore zit vaak een harpoenachtig orgaan aan een lange, aan de voorzijde opgerolde draad, het filament. De achterzijde van het filament is omgeven door een "polaroplast", bestaande uit langwerpige membranen. Onder het filament ligt aan de voorzijde een vesikel.
De manier van voortplanting varieert van soort tot soort. Sommige soorten hebben aseksuele voortplanting, terwijl anderen ook seksuele voortplanting kennen.[2]
De levenscyclus verschilt sterk van soort tot soort. De sporen zijn meestal goed bestand tegen moeilijke omstandigheden en kunnen een aantal jaar buiten een gastheer overleven. Wanneer de spore een mogelijke gastheer bereikt, zwelt het vesikel aan de voorzijde op zodat het filament de inhoud van de spore in het cytoplasma van de gastheer spuit. Binnen in de gastcel groeit daarna een plasmodium, dat zich uitbreidt om ten slotte nieuwe sporen te produceren. Het plasmodium kan zich door de gastheer verspreiden en meerdere organen infecteren.
Soms heeft infectie een dodelijke afloop voor de gastheer. In sommige gevallen kan de stofwisseling en reproductie van de gastcel compleet door de parasiet zijn overgenomen, waardoor een xenoma groeit. Microsporidia zorgen meestal echter voor een chronisch, verzwakkend ziektebeeld in plaats van dodelijke afloop. Bij de gastheer kan infectie zich uiten door verminderde vruchtbaarheid, vermoeidheid of verminderd gewicht. Ook kan verticale overerving van moeder op nageslacht voorkomen. Bij insecten kunnen sporen worden meegegeven in de eitjes om te reproduceren in de larven, een manier van voortplanting die transovariaal genoemd wordt.
Vroeger werden Microsporidia gezien als "primitieve" eukaryoten, vooral vanwege het ontbreken van mitochondriën. Ze werden daarom samen met onder andere Parabasalia bij de verouderde groep Archezoa ingedeeld. Recent onderzoek heeft uitgewezen dat deze classificatie onjuist is. Microsporidia blijken juist verder ontwikkeld te zijn, de afwezigheid van structuren als organellen is te wijten aan het feit dat ze later in de evolutie deze structuren verloren, omdat deze overbodig geworden waren.[3] Bovendien is de voortplantingswijze van Microsporidia ingewikkeld.
Tegenwoordig worden Microsporidia ofwel tot de schimmels gerekend, of gezien als een zustergroep daarvan.[4]
BronnenDe Microsporidia of Microspora vormen een stam van eencellige parasieten, nauw verwant aan schimmels. Er bestaan waarschijnlijk meer dan een miljoen soorten, waarvan er ongeveer 1500 zijn beschreven. Microsporidia leven als parasieten op dieren, voornamelijk insecten. Ongeveer een tiende van alle soorten infecteert gewervelden, waaronder mensen. Sommige soorten worden gebruikt om insectenplagen tegen te gaan.
Infectie vindt plaats door middel van sporen, die de cellen van de gastheer binnendringen. Deze sporen zijn maar 1 tot 40 micrometer groot en behoren daarmee tot de kleinste eukaryotische cellen. Microsporidia vormen daarnaast de groep met het kleinste genoom van alle eukaryoten.
Mikrosporydia (Microsporidia Balbiani, sporowce[1]) – grupa eukariotów o niejasnej pozycji filogenetycznej.
Mikrosporydia są silnie wyspecjalizowanymi organizmami jednokomórkowymi. Nie mają mitochondriów i peroksysomów[2], ich rRNA ma pewne cechy prokariotyczne i wiele delecji, a ich genomy są najmniejsze spośród wszystkich eukariotów[3], wielkością odpowiadają genomom bakterii[4]. Cechy te utrudniają poznanie ich pozycji filogenetycznej – początkowo zaliczano je do protistów, jednak późniejsze badania molekularne sugerowały ich przynależność do grzybów[2][4], w których nadaje się im przeważnie rangę typu[5]. Niektóre analizy molekularne grupują jednak Microsporidia z innymi eukariotami, takimi jak Cryptophyta, Cercozoa, Heterolobosea, Parabasalidea, Arcillenida lub Haplosporidia, lecz nie z grzybami[3].
Są obligatoryjnymi wewnątrzkomórkowymi pasożytami innych eukariotów[2]. Infekują żywiciela z dużą prędkością wbijając w niego specjalną rurkę, przez którą dostają się do wnętrza komórki[4]. Nosema bombycis pasożytuje na jedwabnikach, wywołując u nich tzw. pebrynę. Komórki mikrosporydiów często mają dwa jądra komórkowe i są okryte chityną.
Mikrosporydia (Microsporidia Balbiani, sporowce) – grupa eukariotów o niejasnej pozycji filogenetycznej.
Mikrosporydia są silnie wyspecjalizowanymi organizmami jednokomórkowymi. Nie mają mitochondriów i peroksysomów, ich rRNA ma pewne cechy prokariotyczne i wiele delecji, a ich genomy są najmniejsze spośród wszystkich eukariotów, wielkością odpowiadają genomom bakterii. Cechy te utrudniają poznanie ich pozycji filogenetycznej – początkowo zaliczano je do protistów, jednak późniejsze badania molekularne sugerowały ich przynależność do grzybów, w których nadaje się im przeważnie rangę typu. Niektóre analizy molekularne grupują jednak Microsporidia z innymi eukariotami, takimi jak Cryptophyta, Cercozoa, Heterolobosea, Parabasalidea, Arcillenida lub Haplosporidia, lecz nie z grzybami.
Są obligatoryjnymi wewnątrzkomórkowymi pasożytami innych eukariotów. Infekują żywiciela z dużą prędkością wbijając w niego specjalną rurkę, przez którą dostają się do wnętrza komórki. Nosema bombycis pasożytuje na jedwabnikach, wywołując u nich tzw. pebrynę. Komórki mikrosporydiów często mają dwa jądra komórkowe i są okryte chityną.
Microsporidia é um filo de fungos que sofreu algumas modificações taxonômicas recentemente. Inicialmente, era um grupo de parasitas unicelulares (de animais) e formadores de esporos. No passado, eram incluídos entre os protozoários, mas após estudos genéticos descobriu-se que eram relacionados aos fungos, principalmente aos fungos do antigo filo Zygomycota.[1]. Quando reunidos, criou-se o "novo" filo Microsporidia.
Microsporidia é um filo de fungos que sofreu algumas modificações taxonômicas recentemente. Inicialmente, era um grupo de parasitas unicelulares (de animais) e formadores de esporos. No passado, eram incluídos entre os protozoários, mas após estudos genéticos descobriu-se que eram relacionados aos fungos, principalmente aos fungos do antigo filo Zygomycota.. Quando reunidos, criou-se o "novo" filo Microsporidia.
Microspora. Şubenin tüm elemanları, zorunlu hücre içi parazitleridir. Mitokondrileri bulunmaz ve tek hücreli sporlar meydana getirirler. Böcekler için patojen olan canlıların çoğu, bu şubenin üyeleridir. Hayat evrelerinde hem hücre içi hem de hücre dışı yaşam görülür. Hücre içi eşeyli ve eşeysiz devreler boyunca, konak canlıya zarar verilir. Sonuçta da, çok dayanıklı bir spor hücresi üretilir.
Microspora. Şubenin tüm elemanları, zorunlu hücre içi parazitleridir. Mitokondrileri bulunmaz ve tek hücreli sporlar meydana getirirler. Böcekler için patojen olan canlıların çoğu, bu şubenin üyeleridir. Hayat evrelerinde hem hücre içi hem de hücre dışı yaşam görülür. Hücre içi eşeyli ve eşeysiz devreler boyunca, konak canlıya zarar verilir. Sonuçta da, çok dayanıklı bir spor hücresi üretilir.
У мікроспоридій відсутні мітохондрії але наявні мітосоми. Також відсутні структури, необхідні для руху, джгутики. Високовитривалі спори мікроспоридій здатні виживати зовні організму хазяїна протягом кількох років. Елементи морфології спор є таксономічною ознакою. Спори більшості видів є овальними або грушовидними, але також існують паличковидні та сферичні. Окремі роди мають спори унікальної форми.
Спори мають захисну оболонку, що складається із трьох шарів:
У більшості випадків існує два тісно пов'язаних ядра, що утворюють діплокаріон; іноді є тільки одне ядро. Передня половина спори містить гарпун-подібний апарат із довгими ниткоподібними полярними нитки, які звернуті у задній половині спори. Передня частина полярних ниток оточена поляропластом і мембранними ламелами. За полярними нитка розташована задня вакуоль.[1]
Захворювання, що викликаються мікроспоридіями, несуть як негативні, так і позитивні наслідки для людини. Майже 150 років відзначаються значні збитки у господарстві в наслідок епізоотий нозематозів у тутового Bombyx mori і дубового Antheraea perny шовкопрядів, а також у бджіл Apis mellifera. Крім того описані випадки масової загибелі інших безхребетних: комах-ентомофагів, опилювачів; ракоподібних — рака річкового (Astacus astacus) та крабів; молюсків — мідії їстівної (Mytilus edulus) в марикультурі. Широко поширені мікроспоридіози у риб, заражаючи у них мускулатуру (Pleistophora), мезентерій і кишечник (Glugea), репродуктивні органи (Glugea, Coccospora) і навіть центральну нервову систему (Spraguea lophii). Описані випадки масового зараження (до 70-100%) мікроспоридією Glugea takedai ікринок у горбуши (Oncorhynchus gorbuscha) і сими (O. masou), що призводять до різкого падіння плодовитості цих риб. Випадки захворювання мікроспоридіозами ссавців і птахів як правило закінчуються загибеллю. Особливу проблему являють собою мікроспоридіози людини. До недавнього часу були відомі лише окремі випадки захворювання людей, зазвичай у тих, хто працював на кролячих фермах (паразит кролів — Encephalitozoon cuniculi). На даний час відомо 13 видів паразитів, що викликають у людей, хворих на СНІД, захворювання очей — кератит і блефарит, хронічну діарею або генералізовану міопатію з летальним кінцем.[4]
Протягом певного часу мікроспоридій вважали дуже примітивними еукаріотами, особливо через відсутність мітохондрій, і поміщали разом з іншими найпростішими, такими як Diplomonadida, Parabasalia і Archamoebae до примітивної групи Archezoa. Пізніші дослідження спростували ці положення. Тим не менш, мікроспоридії вважаються дуже розвиненими і спеціалізованими організмами, які просто позбавилися непотрібних функцій, оскільки вони живляться за рахунок хазяїна.[5] В цілому, спороутворюючі організми мають складну систему розмноження, як статевого так і безстатевого.
В даний час мікроспоридій відносять до грибів, або як сестринську групу грибів із загальним предком.[6][7][8][9]
Поділяються на два класи із такою систематикою:
Натомість Воссбрінк і Дебрюнер-Воссбрінк (Vossbrinck & Debrunner-Vossbrinck) запропоновали три класи мікроспоридій, на основі середовища їх існування: Aquasporidia, Marinosporidia і Terresporidia.[10]
Microsporidia là một ngành của giới Nấm. Ngành nấm này từng được coi là động vật nguyên sinh hay sinh vật nguyên sinh, nhưng giờ đây đã được xếp chung với giới Nấm,[6] hoặc nhóm sinh vật có quan hệ gần với nấm.[7]
Dữ liệu liên quan tới Microsporidia tại Wikispecies
Microsporidia là một ngành của giới Nấm. Ngành nấm này từng được coi là động vật nguyên sinh hay sinh vật nguyên sinh, nhưng giờ đây đã được xếp chung với giới Nấm, hoặc nhóm sinh vật có quan hệ gần với nấm.
Оболочка споры у подавляющего большинства видов микроспоридий состоит из трёх слоёв: гликопротеиновой экзоспоры, хитиновой эндоспоры и цитоплазматической мембраны. Экзоспора может быть многослойной и формировать придатки различной формы[4].
Аппарат экструзии включает в себя заднюю вакуоль, якорный диск, поляропласт и полярную трубку. Задняя вакуоль представлена одной или несколькими камерами, расположенными терминально. Иногда внутри вакуоли бывает расположена постеросома — рудимент аппарата Гольджи. Во время экструзии споры данная органелла резко увеличивается в размере, выталкивая зародыш в полярную трубку[5]>.
Поляропласт чаще всего представляет собой упаковку плотно уложенных мембран, но также может включать различные везикулы и трубчатые структуры. Считается, что поляропласт задействован в создании высокого внутриспорового давления, необходимого для выворачивания полярной трубки, а также является «поставщиком мембран» для самой трубки и проходящей по ней спороплазмы[5].
Полярная трубка представлена удлинённой двумембранной структурой с умеренно электроноплотным содержимым, отходящей от расположенного на переднем конце споры якорного диска и уложенной спирально. Длина, степень развития и форма полярных трубок у микроспоридий широко варьируют. Обычно, число витков спирали колеблется от 6 до 12, но у некоторых видов достигает 36. Представители семейства Metchnikovellidae — паразиты других одноклеточных организмов (тоже паразитических) грегарин имеют крайне редуцированный аппарат экструзии, с полярной трубкой по размерам не превышающей длину споры. По особенностям строения выделяют несколько типов полярных трубок: изофилярная — обладает одинаковым размером на всём протяжении; анизофилярная — диаметр первых витков значительно превышает диаметр последующих; гетерофилярная — различные витки имеют различный диаметр. После выброса полярная трубка становится полой, и через неё происходит внедрение спороплазмы в клетку хозяина[5].
Спороплазма (зародыш) представляет собой лежащее в споре одиночное ядро или диплокарион, окружённое тонким слоем цитоплазмы с рибосомами. После экструзии споры зародыш проходит через полярную трубку и оказывается в цитоплазме хозяина. При этом в заражённой клетке спороплазма оказывается окружённой собственной цитоплазматической мембраной, происхождение которой до сих пор доподлинно не известно[6].
Пролиферативная стадия — меронт — представлена интенсивно растущей и размножающейся клеткой, одноядерной или многоядерной, в зависимости от стадии мерогонии[en]. Цитоплазма меронтов содержит минимальный набор органелл: аппарат Гольджи трубчатого типа, строение которого уникально для микроспоридий[7], рибосомы, гладкий и шероховатый эндоплазматический ретикулум. Кроме того, в цитоплазме меронтов можно найти различные трубчатые структуры и везикулы[8].
При переходе к первой стадии спорогонии — споронту — клетка микроспоридии приобретает поверх цитоплазматической мембраны дополнительную оболочку, первое время заметную только при электронном микроскопировании. На этом этапе продолжается деление ядер, в результате чего образуются многоядерные спорогональные плазмодии, дающие начало споробластам.
У многих видов микроспоридий снаружи от оболочки споронта закладывается ещё один покровный слой, образующий оболочку спорофорного пузырька (по старой терминологии панспоробласта). Строение спорофорных пузырьков и развитие полости между оболочками широко варьирует у разных видов микроспоридий[9].
Споробласт — переходная стадия между спорогональным плазмодием и спорой. Характеризуется интенсивным развитием оболочек и закладкой всех будущих органелл споры[9].
Геном микроспоридий, как и у большинства других эукариот, состоит из набора линейных хромосом, но является, однако, крайне редуцированным и необычным. Средний размер геномов микроспоридий чрезвычайно мал, у Encephalitozoon intestinalis он составляет всего 2,3 миллиона пар нуклеотидов, что даже меньше, чем у некоторых бактерий. Набор кодируемых белков также невелик, в геноме Encephalitozoon cuniculi[en] найдено около 2000 белоккодирующих последовательностей. Геном микроспоридий сильно компактизирован, и в нём практически отсутствуют интроны, при этом гены, гомологичные генам других организмов, у микроспоридий часто бывают значительно короче[3].
В процессе адаптации к внутриклеточному паразитизму микроспоридии утратили большинство метаболических путей, таких как окислительное фосфорилирование или цикл Кребса. На стадии внутриклеточного развития паразит вообще не использует собственную систему энергетического метаболизма, полностью полагаясь на транспорт АТФ из цитоплазмы клетки хозяина за счёт уникальных АТФ-переносчиков[10]. Данные переносчики пластидно-бактериального типа были приобретены микроспоридиями за счёт горизонтального переноса генов от бактерий и являются уникальными для данного таксона. Таким образом, микроспоридии дальше всех других эукариот продвинулись на пути адаптации к внутриклеточному паразитизму, так как кроме них пока не было найдено паразитов, способных напрямую транспортировать АТФ из хозяйской клетки[10].
На стадии споры микроспоридии поддерживают свою жизнедеятельность за счёт гликолиза. При этом, долгое время было неясно, каким образом происходит реокисление образующихся в ходе гликолиза восстановительных эквивалентов, таких как NADH. На сегодняшний день предлагается следующая схема данного процесса, включающая глицеролфосфатный челночный механизм[en] и альтернативную оксидазу. В цитоплазме цитоплазматическая форма глицерол-3-фосфатдегидрогеназы окисляет NADH до NAD+, восстанавливая при этом дигидроксиацетонфосфат до глицерол-3-фосфата. Последний поступает в межмембранное пространство митосомы, где вновь окисляется до дигидроксиацетонфосфата за счёт FAD-зависимой митосомальной формы глицерол-3-фосфатдегидрогеназы. Восстановленная форма FADH2, в свою очередь, передаёт электроны на пул убихинонов, расположенный во внутренней мембране митосомы. Конечным этапом является реокисление убихинона ферментом альтернативная оксидаза с использованием кислорода в качестве конечного акцептора электронов и восстановлением последнего до воды[11]. Важно отметить, что в геномах представителей группы Terresporidia (включает в себя большинство микроспоридий, паразитирующих в наземных животных) гена альтернативной оксидазы найдено не было. Таким образом, схема энергетического обмена у этой группы до сих пор остаётся загадкой[11].
Инвазионной стадией у микроспоридий является спора. Она содержит зародыш (спороплазму) и сложно организованный аппарат экструзии, обеспечивающий выброс зародыша из споры и внедрение его в клетку хозяина. Для многоклеточных животных заражение обычно происходит при попадании спор паразитов в пищеварительный тракт хозяина. В отличие от других внутриклеточных паразитов, в подавляющем большинстве случаев развитие микроспоридий происходит в прямом контакте с цитоплазмой клетки хозяина, без образования паразитофорной вакуоли[12]. После внедрения происходит рост и дифференциация зародыша, после чего клетка паразита превращается в меронт, пролиферативную стадию, которая размножается бинарным или множественным делением по типу мерогонии[en]. Затем микроспоридии приступают к спорогонии. Ядра споронтов претерпевают от одного до нескольких делений, в результате чего образуются спорогональные плазмодии, дающие начало споробластам, в которых происходит формирование спор. Обычно на этом этапе происходит разрушение заражённой клетки хозяина, а вышедшие таким образом споры заражают другие клетки того же организма или выводятся наружу для заражения других особей.
Микроспоридии демонстрируют очень высокую степень разнообразия жизненных циклов. Описанный выше моноксенный (включающий только одного хозяина) цикл с образованием спор одного типа характерен для почти 80 % описанных видов[13]. Однако и в таком цикле особенности спорогонии, строение ядерного аппарата, количество и типы делений на всех стадиях широко варьируют. В остальных случаях цикл может проходить более чем в одном хозяине, включать несколько спорогоний с образованием нескольких типов спор, различающихся морфологически и функционально. Для нескольких видов на различных стадиях развития описан мейоз и половой процесс, проходящий по типу гаметоцитогамии[12]. Примером наиболее сложных поликсенных жизненных циклов, включающих половой процесс и морфо-функциональную дифференциацию спор, являются жизненные циклы микроспоридий родов Amblyospora и Parathelonahia.
С момента первого описания в 1857 году положение таксона в системе живой природы постоянно менялось. Первоначально микроспоридии были отнесены к группе Schizomycetes, куда входили различные виды дрожжей и бактерий. Затем, в начале XX века таксон был вместе с другими внутриклеточными паразитами включен в состав группы Sporozoa в подгруппу Cnidosporidia[en], включающую также миксоспоридий, актиноспоридий и хеликоспоридий. Во второй половине XX века было накоплено достаточно данных, доказывающих искусственность группы Sporozoa, и микроспоридии были включены в группировку Archezoa[en], объединяющую одноклеточных организмов без митохондрий. Предполагалось, что представители данной группы являются наиболее древними из современных эукариот и отделились от их общего предка ещё до приобретения митохондрий. Несмотря на то, что сейчас можно считать доказанными ошибочность гипотезы Archezoa и вторичность потери микроспоридиями митохондрий, часть молекулярных исследований, выполненных в конце XX века, поддерживали данную гипотезу[3]. Однако, после увеличения числа исследуемых генов, все больше молекулярно-филогенетических исследований демонстрировали родство микроспоридий с грибами. При этом в разных работах положение микроспоридий относительно грибов оказывается различным. По результатам некоторых исследований микроспоридии входят в состав грибов[15], однако последние работы обычно выявляют микроспоридий, как сестринскую грибам группу[14][16]. На сегодняшний день принято формальное включение микроспоридий в таксон Fungi, однако такая точка зрения подвергается критике со стороны многих специалистов[2]., и вполне вероятно, что систематическое положение микроспоридий ещё не раз кардинально изменится.
Классификация микроспоридий подвергается непрерывным перестройкам на основе использования новых таксономических признаков, полученных при изучении тонкого строения, жизненных циклов, цитохимических и молекулярно-биологических свойств как новых, так и известных форм. В классической системе, построенной на морфологических признаках, общепризнано разделение типа на два класса, один из которых включает формы микроспоридий, образующие в конце коротких жизненных циклов споры с примитивным аппаратом экструзии, а другой — более сложно организованные формы с моно- или диксенными жизненными циклами, завершающимися образованием одного-двух-трёх типов спор с хорошо развитым аппаратом экструзии[17]. При подразделении на отряды и семейства основное внимание уделено строению, в основном, стадий спорогонии, типу ядерного аппарата, хромосомным и жизненным циклам. Однако анализ нуклеотидных последовательностей рибосомальной РНК показывает отсутствие связи между классическими признаками морфологии и филогенетическим родством между видами микроспоридий, что требует пересмотра значения морфологических критериев, используемых в систематике. Использование подходов молекулярной филогении позволяет разделить проанализированные формы микроспоридий на пять филогенетических ветвей, объединяемых в три класса, исходя из первичной приуроченности паразитов к месту обитания хозяев[18].
Микроспоридии вызывают хозяйственные потери на шелководнях и пасеках, будучи возбудителями эпизоотий нозематозов у медоносных пчёл (Apis mellifera), тутового шелкопряда (Bombyx mori). Описаны случаи массовой гибели других полезных беспозвоночных животных, например, мидий Mytilus edulus, а также различных промысловых видов рыб[19].
Особую проблему представляют микроспоридиозы человека. Обычно микроспоридии вызывают тяжелые и быстро развивающиеся оппортунистические инфекции, возникающие на фоне иммунодефицита[20]. Однако данные патогены могут быть опасны и для иммунокомпетентных людей; было показано, что микроспоридия Encephalitozoon cuniculi, паразитирующая у кроликов, способна заразить человека с нормальной иммунной системой[19]. Клинические проявления микроспоридиоза очень разнообразны и варьируют в зависимости от вида микроспоридии-возбудителя инфекции. Наиболее часто встречаемым симптомом является диарея[20].
Микроспоридии, паразитирующие у насекомых-вредителей сельскохозяйственного, медицинского и ветеринарного значения, являются перспективными агентами биологической борьбы с этими видами. Некоторые виды микроспоридий комаров вызывают высокую смертность личинок, снижают плодовитость самок, а также предотвращают развитие нескольких видов малярийных плазмодиев[12]. В 1978 году в США для борьбы с прямокрылыми вредителями был создан первый промышленный биопрепарат на основе микроспоридий — «Нолок», который получил высокую экономическую оценку благодаря низкой стоимости, высокой эффективности и отсутствию негативного влияния на окружающую среду[12].
Оболочка споры у подавляющего большинства видов микроспоридий состоит из трёх слоёв: гликопротеиновой экзоспоры, хитиновой эндоспоры и цитоплазматической мембраны. Экзоспора может быть многослойной и формировать придатки различной формы.
Аппарат экструзии включает в себя заднюю вакуоль, якорный диск, поляропласт и полярную трубку. Задняя вакуоль представлена одной или несколькими камерами, расположенными терминально. Иногда внутри вакуоли бывает расположена постеросома — рудимент аппарата Гольджи. Во время экструзии споры данная органелла резко увеличивается в размере, выталкивая зародыш в полярную трубку>.
Поляропласт чаще всего представляет собой упаковку плотно уложенных мембран, но также может включать различные везикулы и трубчатые структуры. Считается, что поляропласт задействован в создании высокого внутриспорового давления, необходимого для выворачивания полярной трубки, а также является «поставщиком мембран» для самой трубки и проходящей по ней спороплазмы.
Полярная трубка представлена удлинённой двумембранной структурой с умеренно электроноплотным содержимым, отходящей от расположенного на переднем конце споры якорного диска и уложенной спирально. Длина, степень развития и форма полярных трубок у микроспоридий широко варьируют. Обычно, число витков спирали колеблется от 6 до 12, но у некоторых видов достигает 36. Представители семейства Metchnikovellidae — паразиты других одноклеточных организмов (тоже паразитических) грегарин имеют крайне редуцированный аппарат экструзии, с полярной трубкой по размерам не превышающей длину споры. По особенностям строения выделяют несколько типов полярных трубок: изофилярная — обладает одинаковым размером на всём протяжении; анизофилярная — диаметр первых витков значительно превышает диаметр последующих; гетерофилярная — различные витки имеют различный диаметр. После выброса полярная трубка становится полой, и через неё происходит внедрение спороплазмы в клетку хозяина.
Строение спороплазмы и стадий внутриклеточного развитияСпороплазма (зародыш) представляет собой лежащее в споре одиночное ядро или диплокарион, окружённое тонким слоем цитоплазмы с рибосомами. После экструзии споры зародыш проходит через полярную трубку и оказывается в цитоплазме хозяина. При этом в заражённой клетке спороплазма оказывается окружённой собственной цитоплазматической мембраной, происхождение которой до сих пор доподлинно не известно.
Пролиферативная стадия — меронт — представлена интенсивно растущей и размножающейся клеткой, одноядерной или многоядерной, в зависимости от стадии мерогонии[en]. Цитоплазма меронтов содержит минимальный набор органелл: аппарат Гольджи трубчатого типа, строение которого уникально для микроспоридий, рибосомы, гладкий и шероховатый эндоплазматический ретикулум. Кроме того, в цитоплазме меронтов можно найти различные трубчатые структуры и везикулы.
При переходе к первой стадии спорогонии — споронту — клетка микроспоридии приобретает поверх цитоплазматической мембраны дополнительную оболочку, первое время заметную только при электронном микроскопировании. На этом этапе продолжается деление ядер, в результате чего образуются многоядерные спорогональные плазмодии, дающие начало споробластам.
У многих видов микроспоридий снаружи от оболочки споронта закладывается ещё один покровный слой, образующий оболочку спорофорного пузырька (по старой терминологии панспоробласта). Строение спорофорных пузырьков и развитие полости между оболочками широко варьирует у разных видов микроспоридий.
Споробласт — переходная стадия между спорогональным плазмодием и спорой. Характеризуется интенсивным развитием оболочек и закладкой всех будущих органелл споры.
微孢子蟲(學名:Microsporidea)為羅茲菌門下的一綱。它是由孢子形成的單細胞寄生蟲。目前多於一百萬種微孢子蟲中的1500種版命名。微孢子蟲只能寄生於動物宿主。大部份的動物物種都可以被微孢子蟲寄生,包括人類。它多數感染昆蟲,也是甲殼和魚的常見疾病。一般的来说,已经命名的微孢子蟲會專門感染指定的物種。而部分物種可能會感染人類,它們多是伺機感染原,在人體免疫力下降時才造成感染。
約有10%的物種是脊椎動物的病原。

微胞子虫(びほうしちゅう、Microsporidia)はさまざまな動物の細胞内に寄生する単細胞真核生物の一群で、これまでに1200種以上が知られている。昆虫、甲殻類、魚類、ヒトを含む哺乳類などに感染する病原体が多く含まれている。かつては粘液胞子虫(現在は多細胞動物起源とされる)とともに原生動物門胞子虫綱、あるいは極嚢胞子虫綱に入れられたが、現在では極めて特殊化した菌類だと考えられ、独立な分類群と考えられている。
微胞子虫の胞子はたいてい卵形をしていて1-40 μm程度の大きさであり、タンパク質とキチン質からなる2層の細胞壁によって保護されている。胞子の内部には特徴的なコイル状の構造があり、これはミクソゾア類の極糸と混同しないよう極管(polar tube)と呼ばれている。微胞子虫類にはミトコンドリアがなく、おそらく相同なマイトソーム(mitosome)を持っている。菌類と共通した特徴として、鞭毛のような運動器官がないこと、2つの細胞核が細胞内で密接する複核(diplokaryon)が存在することが多いことが挙げられる。
微胞子虫の生活環はかなり多様である。無性生殖のみによる単純な生活環を持つものもあれば、複数の宿主で無性生殖と有性生殖が交替する複雑な生活環を持つものもある。感染の初期は個体内で増殖するための胞子を作り、その後個体間で伝播するための胞子を作るような種もある。
通常は経口感染によって水平伝播するが、昆虫宿主の場合にはしばしば宿主の卵巣から卵に渡される経卵巣感染によって垂直伝播が起きる。極管の射出の引き金は種によって様々なものが知られているが、例えば高pHや、ポリグルタミンやムチンの存在などがある。この例では消化管の粘膜の近くで条件が満たされ、胞子の内圧が上がり、極管がホースを裏返しにするようにして宿主細胞に突き刺さる。このとき極管は数百μmと長く伸びているが、胞子の中の胞子原形質(sporoplasm)は極管を通って数十秒という短時間で宿主細胞に注入される。また宿主細胞に貪食されることでも感染が成立する。
胞子原形質は宿主細胞の中で成長し、細胞分裂を繰り返したり多核の変形体になったり種によって様々である。感染細胞は代謝活性が上がったり、肥大化して表面積の大きな腫瘍を形成したりと、多様な変化を示すが、これによって微胞子虫の栄養体に充分な栄養が供給されるようになる。その後胞子を形成するが、この時に有性生殖が見られる場合がある。
宿主に対する影響は、寿命、生殖能力、体重及び一般的な体力の減少などがある。したがって致死的というよりむしろ慢性的で衰弱させてゆくような病気を引き起こすことが多い。 昆虫、甲殻類、魚類に寄生して農業・漁業に深刻な影響を与える病原体が知られる。 また哺乳類でも自然治癒性の下痢の病原体としてごくありふれているほか、日和見感染を起こす病原体が知られている。 このほか、粘液胞子虫や、グレガリナ類などに寄生する微胞子虫が知られている。
1950年代までは微胞子虫は人間には病原性を示さないと考えられていた。人体に感染することが知られたのは1959年が初めてであり、その後もしばらくの間はほとんど報告がなかった。しかしこれは健康な成人に感染しても自然治癒するか重い症状がないため気付かれなかっただけで、罹患率は人口の2%ほど、報告によっては70%に達するケースもあるほどありふれた病原体である。乳児や、臓器移植やAIDSなどで免疫力が低下した状態では重篤な症状になることがあり、日和見感染症として注目されるようになってきている。
人間への感染源としては、家畜、家禽、ペットの鳥や犬などが主であり、人獣共通感染症である。動物保護施設では3割の犬が感染していたことがあり、子犬の集団死の原因にもなる。主に経口感染によると考えられるため、飲料水が感染源になっている可能性もある。種によっては性感染や呼吸器からの感染の可能性もある。動物では先天性感染が知られているが、これまでのところ人間での報告はない。角膜への外傷性感染も知られている。感染部位は消化管が最も多いが、生殖器、呼吸器、排泄器、神経系などにも感染するものがある。
人体病原体としては8属13種が知られている。代表的なものについて症状を下に示したが、この他に、Trachipleistophora, Pleistophora, Nosema, Vittaforma, Microsporidiumなどが間質性角膜炎や筋炎を起こすことが知られている。
微胞子虫類は真核生物としては最も小さいゲノムを持つ生物群である。これまで知られている限りでは1000-2000万塩基対のものが多いが、Encephalitozoon属では極めて小さく300万塩基対に満たない。これは原核生物である大腸菌よりも小さい。
2001年にEncephalitozoon cuniculiの全ゲノム配列が解読され、極端に濃縮されたゲノム構造が明らかになった。E. cuniculiのゲノムは11本の染色体からなり、1本の染色体は22-32万塩基対というごく小さなものである。中でも全ての染色体の両端にリボソームRNA遺伝子が並んでいることは注目すべき特徴である。二次共生藻類のヌクレオモルフは真核生物の核が極端に縮小されたものだと考えられるが、クリプト藻とクロララクニオン藻という系統的に無関係な藻類の両方が、やはりヌクレオモルフの全ての染色体の両端にリボソームRNA遺伝子を持っている。したがって、極端に小さなゲノムとリボソームRNA遺伝子の配置の間にはなにか因果関係があるだろうと考えられている。
微胞子虫類は単細胞であることから、伝統的に原生動物とみなされてきた。そして、他のミトコンドリアを欠く真核生物と同様に非常に古い時期に分岐した最も原始的な真核生物だと考えられたこともある。しかし、最近の分子系統解析や生化学的傍証によれば、微胞子虫は接合菌などに近縁な菌類であり、寄生生活に適応してミトコンドリアが退化したものと考えられている。
微胞子虫類は現在では独立の門として扱うことが多いが、その学名が何であるかについては混乱が見受けられる。1980年の国際原生動物学会による合意体系には微胞子虫門(phylum Microspora Sprague, 1977)とあるため、これを正式なものと位置付ける研究者は多いが、そうする研究者でさえ通常はmicrosporidiaという名を好んで使う。これはかつて亜綱をあてていた頃の名称であるが、phylum Microsporidiaと表記する研究者もいる。2005年の合意体系では「門」などの階級名称そのものを放棄しているが、微胞子虫類はMicrosporidia Balbiani, 1882とされている。ちなみに緑藻にMicrospora属という生物がある。
現在のところ核の存在様式に注目した分類体系(Sprague et al. 1992)が用いられているが、核の存在様式は簡単に変化しうる特徴であり生物の系統をうまく反映しないことがわかっている。最近になって分子系統解析に基づき宿主の分布域(淡水産・海水産・陸上産)に注目するとうまくいきそうだという提案(Vossbrinck and Debrunner-Vossbrinck 2005)が出ているが、今後この方向性が受け入れられるかどうかは定かでない。また属や種のレベルでの分類も分子系統解析によって盛んに整理され始めたところである。
微胞子虫(びほうしちゅう、Microsporidia)はさまざまな動物の細胞内に寄生する単細胞真核生物の一群で、これまでに1200種以上が知られている。昆虫、甲殻類、魚類、ヒトを含む哺乳類などに感染する病原体が多く含まれている。かつては粘液胞子虫(現在は多細胞動物起源とされる)とともに原生動物門胞子虫綱、あるいは極嚢胞子虫綱に入れられたが、現在では極めて特殊化した菌類だと考えられ、独立な分類群と考えられている。
미포자충류(微胞子蟲類)는 다양한 동물의 세포 내에 기생하는 단세포 진핵생물의 한 무리로, 지금까지 약 1,200종 이상이 알려져 있다. 곤충, 갑각류, 어류, 사람을 포함한 포유류 등을 감염시키는 병원체가 많이 포함되어 있다. 이전에는 점액포자충(현재는 다세포동물 기원으로 보고 있다)과 함께 원생동물문의 포자충강 또는 극낭포자충강으로 분류했지만, 현재는 지극히 특수한 균류로 생각하고 있으며, 독립적인 분류군으로 분류하고 있다.
다음은 후편모생물의 계통 분류이다.[1][2][3][4]
후편모생물 홀로미코타 크리스티디스코이데아 균류/BCG2[5]
BCG1[5]
로젤라아계/Namako-37
홀로조아 플루리포르메아 필로조아 깃편모충류