The type specimen of Paramylodon harlani is a partial right mandible (lower jaw bone) that was found in Big Bone Lick, Boone County, Kentucky (McDonald 1995).
Paramylodon harlani belongs to a family of sloth called Mylodontidae. This family also includes the genera Mylodon and Glossotherium, which are closely related to Paramylodon. It is thought that Paramylodon is a descendant of Glossotherium chapadmalense, a South American sloth that lived during the Blancan period of the Pliocene (4.7-1.8mya) (McDonald 2005; McAfee 2009). This species has characters that are shared by Paramylodon and Glossotherium, but is more similar to the latter genus (McAfee 2009). Its caniniform teeth (canine-like teeth in the front of the jaw) are longer than those of P. harlani and has a wider skull (McAfee 2009).
Paramylodon harlani is a species of giant ground sloth that lived during the Pliocene and Pleistocene epochs in North America (5.3 mya-10 kya). It was a large mammal, about 10 feet (3m) long and weighed about a tonne. They had powerful claws on their hands that they likely used to dig up tubers. They were herbivores and they belong to the family Mylodontidae and are relatives of the South American species, Mylodon darwinii.
Paramylodon harlani was endemic to North America. Its remains have been found in California, Colorado, Oregon, Washington, Montana, and Idaho, in the west, (Stock 1925; Kurten and Anderson 1980; Jefferson 1991; Coltrain et al. 2004; Prothero and Raymond 2011; McDonald 2012b); on the great plains, in Alberta, and Saskatchewan in Canada, North, and South Dakota, Nebraska, Kansas, Oklahoma, Texas, Arizona and New Mexico (Brown 1903; McDonald et al. 2004; McDonald 2012b); and in Kentucky, Florida, Georgia, South Carolina and Pennsylvania on the East Coast (Gidley and Gazin 1938; Roth and Laerm 1980; McDonald 1995; Morgan and Hulbert 1995; Hulbert and Pratt 1998).
Paramylodon harlani was not as large as other ground sloths that lived in North America at that time. It was about 130 cm high at its back (about 4.5 feet) and weighed one tonne on average (McDonald 2005). It had a number of bony scutes or osteoderms underneath its skin (Kurten and Anderson 1980). These functioned as added protection. Its body was covered in shaggy hair. It had a massive chest, powerful forelimbs, stout hind limbs, claws, and a robust tail (Kurten and Anderson 1980). Its body weight was supported on its fifth metatarsal (bones that connect with the toes) and calcaneus (heel bone)(McDonald 2012a). To put it simply, it walked on the outside of its feet and heels. Its skull was elongated and had a blunt nose. The largest skull ever discovered was from Colorado and measured 54 cm in length (1.7 feet) (Kurten and Anderson 1980). It had claws on digits 2, 3 and 4 (Stock 1925). Its neck was short and thick and did not allow for much movement. It might have had thick broad lips, likely prehensile in nature (Stock 1925). When it walked on all four limbs, it’s hands rested on digits 4 and 5 (Stock 1925).
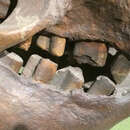
Like other sloths, the cheek teeth of Paramylodon harlani lacked a distinct crown, did not have any distinct roots and grew continuously. It is thus hard to determine the degree of hypsodonty in these sloths. Hypsodonty is a dental condition where animals have large tooth crowns or the part that sticks up above the gum. However, the depth of the mandible or lower jaw bone relative to the length of the alveolar length (length of the row of holes where the teeth insert into the jaw) of the mandibular tooth row can be used to derive an index of hypsodonty. As the height of the teeth increases, the depth of the jaw increases, but the length of the tooth row remains the same. The depth of the jaw is less than the length of the tooth row in less hypsodont sloths while it is more than the length of the tooth row in more hypsodont sloths. Interestingly, in Paramylodon harlani, the ratio of mandibular depth to alveolar tooth row length increases from the Irvingtonian (1.8 mya-240 kya) to the Rancholabrean (240 kya-11.7 kya), suggesting an increase in hypsodonty. This increase correlates with the change in habitats from more closed woodlands, to more open grasslands (McDonald 1995; Bargo et al. 2006). Open grasslands are characterized by coarse vegetation. Increased hypsodonty in the sloths would have helped them process this tough vegetation. They also had caniniform teeth (canine-like), but this trait is highly variable in Paramylodon. These teeth have been found in the Irvingtonian, but some sloths from the Rancholabrean have lost them (McDonald 2005).
These caniniform teeth have different occlusal surfaces (grinding surfaces), and have been associated with two skull morphs, a robust morph that might represent a male and a gracile morph, that might represent the female (McDonald 2006; Prothero and Raymond 2008). In the past, these skull morphs were associated with a subspecies of P. harlani, the robust skull belonging to P. harlani harlani and the gracile skulls belonging to P. harlani tenuiceps (Stock 1925; McDonald 2006). These two morphs become more similar through time, with the Rancholabrean skulls being more similar than Irvingtonian skulls (McDonald 2006).
Paramylodon harlani was a large herbivorous sloth that lived in North America. P. harlani seems to have had a fairly wide thermal niche given its geographic distribution (McDonald 2005). It lived from Canada in the North to Mexico in the South, a region that encompasses a wide range of high and low temperatures. It probably lived alone or in small groups (McDonald and Pelikan 2006). Extant sloths produce one offspring at a time, and it is likely that P. harlani did the same (McDonald and Pelikan 2006).
While there is no direct evidence to infer the diet of P. harlani (no dung has been found), we can look to its cousin, Mylodon darwinii from Chile. Mylodon foraged on grasses and sedges (Moore 1978). This fits with earlier hypotheses that classified P. harlani as a grazer (Stock 1925). Recently, isotopic analyses of P. harlani teeth from Texas have also shown that P. harlani fed primarily on a diet of grasses (Ruez 2005). However, other studies suggest that P. harlani fed on a diet that consisted of leaves as well (Naples 1989). Isotopic studies have also placed P. harlani between ruminants (animals that chew their cud like a cow) and non-ruminants (Coltrain et al. 2004).
Columbian mammoths, Mammuthus columbi, and Paramylodon harlani overlap in their ranges during the Rancholabrean (240 kya- 11.7 kya) (McDonald and Pelikan 2006). These two species fed primarily on grasses and sedges, but do not seem to partition their resources (McDonald and Pelikan 2006). Extant sloths lack a caecum or the first portion of the large intestine (McDonald and Pelikan 2006). Presumably, extinct species lacked one as well. This would have made them foregut fermenters, able to pass food through their digestive system slowly and efficiently, extracting the most out of it (McDonald and Pelikan 2006). Elephants, and mammoths by association, are hindgut fermenters and pass food quickly through their guts (McDonald and Pelikan 2006). This is inefficient and forces them to consume vast quantities of food continuously. McDonald argues that the differences in digestive systems and social structure (mammoths were likely migratory while the sloths were probably territorial) would have allowed them to not compete with each other for food sources (McDonald and Pelikan 2006).
In addition to Mammoths, P. harlani lived with other mammals like camels, antelope, sabertooth cats, dire wolves and bears. While P. harlani probably used claws for defense against Smilodon, Leo atrox, dire wolves, and other predators, it also used them to dig up tubers (Kurten and Anderson 1980; Bargo et al. 2000)
P. harlani at Rancho La Brea, California did not show any changes in size with the changing climates of the late Pleistocene that lasted about 35000 years (Prothero and Raymond 2011). However, other studies have shown that P. harlani increased in size from the Irvingtonian to the Rancholabrean (McDonald 2005).
The taxonomy of Paramylodon harlani has gone through many revisions. These have been described in McDonald (1995), and are summarized here. Richard Owen described two South American genera, Mylodon darwinii and Glossotherium in 1840 (McDonald 1995). In 1842, Owen described another species of Mylodon-M. robustus and in that same publication, suggested that Glossotherium and Mylodon were synonymous (Owen 1842). About 10 years earlier Richard Harlan, an American paleontologist, described sloth remains from Kentucky that he called Megalonyx laqueatus (Harlan 1831), but then redescribed them as a Mylodon in 1835 (Harlan 1835). Owen later gave the Kentucky sloth the species name harlani, after it’s discoverer, Harlan (McDonald 1995). If Owen’s original 1842 synonymization of Mylodon with Glossotherium is to be accepted, then Glossotherium would be the type genus of darwinii, and indeed, Lydekker proposed this reclassification in 1894 (Lydekker 1894). Kraglievish (1928) assumed that Glossotherium had no species assigned to it (because Owen had not assigned one and had simply synonymized Mylodon with it) and classified the bone fragments Owen had described in 1842 as G. uruguayense (Kraglievich 1928). While this reclassification was going on, Barnum Brown described a species of sloth from Nebraska and named it Paramylodon nebrascensis (Brown 1903). Stock (1925) suggested that there were two species of American Mylodonts, M. harlani and P. nebrascensis (Stock 1925); a view that was later modified by Kraglievich (1928) to synonymize harlani and nebrascensis into P. harlani (harlani taking precedence since it was discovered before nebrascensis). Additionally, since the sloth described by Owen-Mylodon-was different from the North American specimens, Kraglievich (1928) assigned the genus Paramylodon to all North American Sloths.