
Beggiatoa és un gènere de bacteris de l'ordre Thiotrichales. Porten el nom en honor del metge i botànic italià F.S. Beggiato. Els organismes viuen en ambients rics en sofre.
Beggiatoa pot habitar tant en aigües dolces com salades, normalment es troba en hàbitats amb alts nivells de sulfur d'hidrogen. Aquests ambients inclouen emanacions fredes del llit marí, fonts de sofre, aigües residuals, capes de fang de llacs i prop de respiradors hidrotermals profunds. Així mateix també es poden trobar a la rizosfera de plantes d'aiguamoll.[1][2]
Les cèl·lules incolores són ovalades o cilíndriques, formant filaments llargs amb un diàmetre de cèl·lules d'entre 12 a 160 micròmetres (segons subespècie). Un vacúol central s'utilitza per a l'acumulació de nitrat, presumiblement per a l'ús d'aquest com a acceptador d'electró en l'oxidació anaeròbica de sulfur.[1]
Els filaments estan envoltats per bava i es poden moure lliscant.
La beggiatoàcia creix de forma quimicoorganoheteròtrofa oxidant composts orgànics a diòxid de carboni en presència d'oxigen, no obtant les altes concentracions d'oxigen poden ser un factor restrictiu. Els composts orgànics són també la font de carboni per a la biosíntesi. Algunes espècies poden oxidar sulfur d'hidrogen a sofre com a font suplemental d'energia (propietat lito-heteròtrofa). El sofre produït s'emmagatzema intracel·lularment.
Algunes espècies tenen l'habilitat de creixement quimicolitoautòtrofa per mitjà d'oxidació de sulfur a energia i amb diòxid de carboni com a font de carboni per a la biosíntesi. En aquest metabolisme el nitrat emmagatzemat és l'acceptador d'electró i reduït a amoníac.
Les espècies autòtrofes marines de Beggiatoàcia són capaces d'oxidar sofre intracel·lular a sulfat.
Un mecanisme freqüentment que es dóna davant la manca d'oxigen, és la reducció de sofre elemental. El sofre es redueix a sulfur consumint el carboni emmagatzemat o per addició de gas d'hidrogen. Possiblement es tracta d'un mecanisme de supervivència davant de períodes sense oxigen.[3]
Els filaments formen estores denses en els sediments d'estuaris, lleixes, filtracions, i respiradors hidrotermals marins profunds. Formant una capa blanquinosa. Donat que són presents i floreixen en ambients marins que han estat subjectes a contaminació, es poden considerar com a espècie indicadora.
Beggiatoacia i uns altres bacteris filamentous relacionats poden provocar problemes si s'instal·len en depuradores, llacunes de residus industrials, etc.
Les beggiatoàcies són capaces de desintoxicar sulfur d'hidrogen a terra.
Beggiatoacia és un parent proper de Thioploca.[1]
Beggiatoa és un gènere de bacteris de l'ordre Thiotrichales. Porten el nom en honor del metge i botànic italià F.S. Beggiato. Els organismes viuen en ambients rics en sofre.
Beggiatoa ist eine Gattung von Gammaproteobakterien, die zur Ordnung Thiotrichales (alias Beggiatoales) gehören. Diese Bakteriengattung war eine der ersten, die der russische Botaniker Sergei Winogradski (englisch auch Winogradsky) entdeckte. Während seiner Forschungen im botanischen Labor von Anton de Bary im Jahr 1887 stellte er fest, dass Beggiatoa Schwefelwasserstoff (H2S) als Energiequelle oxidiert und dabei intrazelluläre Schwefeltröpfchen bildet, wobei Sauerstoff der terminale Elektronenakzeptor ist und CO2 als Kohlenstoffquelle dient. Winogradski bezeichnete diese Form des Stoffwechsels als „Inorgoxidation“ (Oxidation anorganischer Verbindungen), heute Chemolithotrophie genannt. Die Beggiatoa-Arten leben in schwefelreichen Umgebungen auf Böden, sowohl im Süßwasser als auch im Meer, etwa in den hydrothermalen Schloten der Tiefsee und in verschmutzten Meeresumgebungen. Diese Entdeckung war die erste Entdeckung einer Lithotrophie.[2][3] Diese farblosen Bakterien bilden fadenförmige Kolonien (Filamente, wie bei filamentösen Cyanobakterien auch Trichome genannt)[4] und kann in einem Biofilm angeordnet sein, der mit bloßem Auge sichtbar ist und aus sehr langen weißen Fadenmatten besteht; die weiße Farbe ist dabei auf den gespeicherten Schwefel zurückzuführen. Die Beggiatoa-Bakterien sind dabei manchmal mit anderen Schwefelbakterien vergesellschaftet, z. B. mit der Gattung Thiothrix Winogradsky 1888 aus derselben Familie Thiotrichaceae Garrity et al. 2005.[5]
Formell beschrieben wurden zwei Arten von Beggiatoa: die Typusart Beggiatoa alba und Beggiatoa leptomitoformis, wobei letztere erst 2017 veröffentlicht wurde.[6][7] Diese Arten von Beggiatoa haben Zellen mit einem Durchmesser von bis zu 200 µm und gehören damit zu den größten Prokaryoten der Erde.[5]
Wegen des Mangels an Reinkulturen war lange Zeit nur wenig über die Genetik von Beggiatoa bekannt. Beggiatoa alba hat einen GC-Gehalt zwischen 40 und 42,7 mol%. Es gibt zwei oder drei Plasmide; die Genomgröße Stammes Beggiatoa alba B18LD beträgt etwa 3 Mbp (Megabasenpaare).[5] In einer wichtigen Studie über das Genom von Beggiatoa wurden die Sequenzen von zwei Einzelfilamenten eines vakuolisierten Stammes analysiert. Die optische Kartierung zeigte, dass die Genomgröße etwa 7,4 Mbp beträgt, und die Sequenzanalysen zeigten Stoffwechselwege für die Schwefeloxidation, die Nitrat- und Sauerstoffatmung sowie die CO2-Fixierung nach, was die chemolithoautotrophe Physiologie von Beggiatoa bestätigt. Darüber hinaus deutet die vergleichende Genomik auf einen horizontalen Gentransfer (HGT) von Speicher-, Stoffwechsel- und Gleitfähigkeiten zwischen Beggiatoa und Cyanobakterien hin.[8]
Auch das Genom des Stammes Beggiatoa leptomitiformis D-402T (alias DSM 14946) wurde vollständig sequenziert. Die Größe beträgt 4.265.296 bp (Basenpaare), dazu kommt ein Plasmid von 6.185 bp; beide Teile zusammen kodieren nach Vorhersage 3.636 Gene.[9]
Die Gattung Beggiatoa ist insgesamt eine ziemlich vielfältige Gruppe, deren Vertreter verschiedene Lebensräume und Nischen sowohl im Süß- als auch im Salzwasser besiedeln. In der Vergangenheit wurden sie aufgrund ihrer Morphologie und Motilität als Cyanobakterien der Gattung Oscillatoria klassifiziert,[10] aber Analysen der 5S (5S-RNA der ribosomalen RNA) von Stahl et al. (1987) haben gezeigt, dass die Mitglieder von Beggiatoa phylogenetisch weit von Cyanobakterien entfernt sind und zum Stamm der Gammaproteobakterien gehören.[11] Der erwähnte HGT mit Übernahme etlicher Cyanobakterien-Gene dürfte ein wesentlicher Grund für diese Übereinstimmungen und damit für die Unsicherheiten in der Taxonomie sein. Insbesondere ist die Zugehörigkeit weiterer ehemals als Mitglieder der vermeintlichen Cyanobakterien-Gattung Beggiatoa klassifizierten Spezies weiter in der Diskussion, denn diese können womöglich echte Cyanobakterien sein (s. u.).
Nach der List of Prokaryotic names with Standing in Nomenclature (LPSN)[6] und dem National Center for Biotechnology Information (NCBI)[12] sind derzeit (Stand 6. Januar 2022) nur zwei Spezies der Gattung Beggiatoa gültig veröffentlicht worden: Beggiatoa alba und Beggiatoa leptomitoformis.
Die nächsten bekannten Verwandten von Beggiatoa sind Bakterien der Gattung Thioploca (Stand 1999).[13] Die beiden Gattungen Beggiatoa und Thioploca unterscheiden sich durch ein einziges morphologisches Merkmal: Die Thioploca-Filamente treten in Bündeln auf, die von einer gemeinsamen Hülle umgeben sind, während die Beggiatoa-Filamente als Einzelfilamente auftreten ohne diese Struktur auszubilden. Alle Beggiatoa- und Thioploca-Stämme sind in der Lage, Sulfid (bzw. H2S) zu elementarem Schwefel zu oxidieren, der in Form der intrazellulären Schwefelkügelchen gespeichert wird. Dadurch werden die Zellen sehr widerstandsfähig und unter dem Mikroskop auffällig. Diese Eigenschaft, zusammen mit dem Fehlen photosynthetischer Pigmente (Chlorophylle) unterscheidet die Gattungen Beggiatoa und Thioploca als fadenförmige Vertreter der „farblosen Schwefelbakterien“ von anderen fadenförmigen Bakterien, wie den Cyanobakterien und nicht-schwefeloxidierenden Heterotrophen wie Cytophaga und Flexibacter (beide Cytophagaceae, Abteilung Bacteroidetes). Entgegen früherer Auffassung gibt es keine enge evolutionäre Verwandtschaft zwischen Beggiatoa und Thioploca einerseits und anderen gleitenden Fadenbakterien andererseits.[5]
Ein weiteres charakteristisches Merkmal ist die Fähigkeit der weit verbreiteten marinen Arten von Beggiatoa und verwandter Bakterien, Nitrat in den Vakuolen der Zellen zu speichern. Untersuchungen der 16S rRNA-Sequenzen ergaben, dass dieses Merkmal von den Mitgliedern einer monophyletischen Gruppe innerhalb der Gattungen der Beggiatoa-Verwandtschaft geteilt wird. Zu dieser Gruppe gehören auch die Gattungen von Thioploca und Thiomargarita, die sich beide nur geringfügig von Beggiatoa unterscheiden: Während die Zellen von Thioploca in einer gemeinsamen Schleimhülle wachsen, hat Thiomargarita kein fadenförmiges Wachstum erhalten, sondern bildet Ketten aus runden Zellen. Da die überkommene Nomenklatur diese phylogenetische Geschichte nicht widerspiegelt, ist eine neue Benennung der Gattungen und Arten erforderlich.[13] Der neue Referenzstamm (englisch Neo-type strain) für die Süßwasserart Beggiatoa alba ist beispielsweise dann B18LB und dieser bestimmt die Kriterien für die Identifizierung dieser Spezies.[14]
Die Gattung Beggiatoa wird entweder als Mitglied der Familie Beggiatoaceae Migula 1894 (nach LPSN[6]) oder der (etwas größeren) Familie Thiotrichaceae Garrity et al. 2005 (nach NCBI[12] und WoRMS[15]) geführt.
Diese 2005 neu gefasste Familie Thiotrichaceae umfasst nach NCBI nicht nur die Beggiatoaceae, sondern auch die Familien Leucotrichaceae Buchanan 1957 und Achromatiaceae Massart 1901.
In beiden Fällen gilt die Beggiatoa enthaltende Familie als Mitglied der Ordnung Thiotrichales Garrity et al. 2005 Entweder gilt der Ordnungsname Beggiatoales Buchanan 1957 als Synonym von Thiotrichales (nach LPSN[6]), oder die Thiotrichales gelten als die Beggiatoales mit einschließend (nach NCBI[12]).
Mit Beggiatoa verwandte Gattungen in der Familie Beggiatoaceae bzw. Thiotrichaceae sind (Auswahl):
Anm.: AlgaeBase klassifiziert die Gattung Beggiatoa unspezifisch in Taxa der Ränge Familie bis Reich (en. kingdom) Eubacteria.[16]
Quellen der folgenden Artenliste ist LPSN,[6] AlgaeBase,[16] WoRMS,[15] NCBI[12] (U=unsicher, P=provisorisch)
Gattung Beggiatoa V.B.A.Trevisan 1842/1845, mit Schreibvarianten Beggiatoaea, Beggiatonia
Auswahl einiger Neuzuordnungen (Stand 6. Januar 2022):[6][16][15][12]
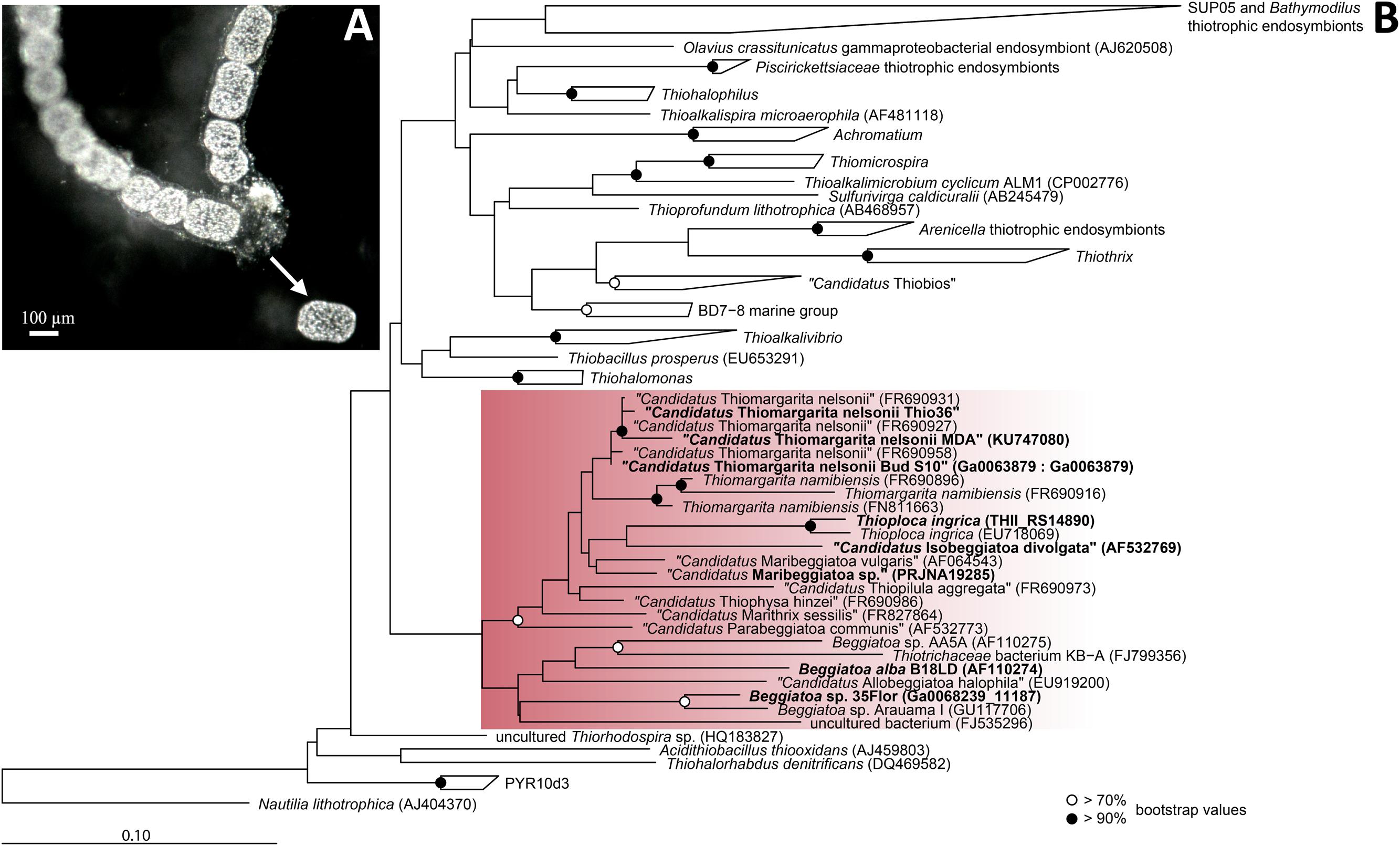
Über diese Neuzuordnungen hinaus erwies sich die Gattung Beggiatoa in den Jahren 2005 bis 2011 als nicht monophyletisch. Um das Problem zu beheben, wurden als neue Kandidatengattungen Isobeggiatoa, Maribeggiatoa, Parabeggiatoa und Allobeggiatoa vorgeschlagen, um einzelne bisherige Beggiatoa-Spezies aufzunehmen. Ein phylogenetischer Baum dieser und weiterer Spezies der Familie Thiotrichaceae findet sich bei der Gattung Thiomargarita §Äußere Systematik.
Winogradski benannte die Gattung zu Ehren von Francesco Secondo Beggiato,[2][3] einem italienischen Arzt und Botaniker aus Vicenza (1806-1883)[29], der die Abhandlung „Delle Terme Euganea“ (Über die Euganeischen Thermen[30][31]) verfasste.[6]
Das Typusart-Epitheton kommt von lateinisch alba ‚die weiße‘.
Anhand ihrer morphologischen Merkmale lassen sich die Beggiatoa-Arten und -Stämme in drei Kategorien einteilen:
Natürlich ist diese Klassifizierung idealisiert, so dass es Ausnahmen geben kann. Schmale Filamente bestehen in der Regel aus länglich-zylindrischen Zellen, deren Länge etwa das 1,5- bis 8-fache ihrer Dicke beträgt, breitere Filamente bestehen hingegen aus eher scheibenförmige Zellen mit Längen, die das 0,10- bis 0,90-fache ihrer Zellbreite betragen. Bei allen kultivierten Stämmen sind die Endzellen der Filamente abgerundet. Obwohl es sich um gramnegative Bakterien handelt, weisen die Beggiatoa-Zellen eine ungewöhnliche Zellwand- und Membranorganisation auf. Manchmal gibt es weitere Membranen, die die Peptidoglykanschicht (Mureinschicht) bedecken, die Anzahl dieser Zusatzmembranen ist sehr unterschiedlich. Ihr Vorhandensein ist möglicherweise auf die rauen Bedingungen zurückzuführen, unter denen einige dieser Organismen leben. Sogar die intrazellulären Granula können von zusätzlichen Membran-Strukturen bedeckt sein. Neben den Schwefelkörnchen finden sich in den Zellen oft auch ähnlich gelagerte Polyhydroxybutyrat- und Polyphosphatkörnchen. Sehr häufig in großen marinen vakuolierten (mit Vakuolen gefüllten) Beggiatoa sind „hohlstrukturierte“ Filamente (en. hollow-structured filaments), die aus schmalen Zellen mit einer großen Zentralen Vakuole zur Speicherung von Nitrat und wenig Zytoplasma bestehen.[5][27]
Die Filamente bewegen sich gleitend, und diese Bewegung ist vermutlich mit fadenartigen Strukturen in der äußeren Membran und trans-Peptidoglykan-Kanälen verbunden. Manchmal können die Filamente auch (ähnlich wie bei den Cyanobakterien der Gattung Oscillatoria) durch die Bildung von Nekridien (Necridia, abgestorbenen Zellen, vgl. Nekrose) in der Mitte des Filaments durchbrechen. Die Motilität der Filamente ist für die Anpassungsfähigkeit der Bakterien von großer Bedeutung. Sie ermöglicht ihnen, sich in Richtung geeigneterer Bedingungen für den Zellstoffwechsel zu bewegen. Die wichtigsten Triebkräfte für die Bewegung der Beggiatoa-Filamente sind ein hoher Sauerstoff- und Sulfidgehalt (O2 und H2S), sowie Lichteinwirkung, von der sich die Filamente wegbewegen (negative Phototaxis).[27]
Die Vermehrung von Beggiatoa geschieht durch Fragmentierung der Filamente (Trichome). Das Wachstum einer Kolonie führt zur Mattenbildung durch abwechselnde Fadenverlängerung und Fadenbruch. Von grundlegender Bedeutung für die Fragmentierung sind (ähnlich wie bei den Cyanobakterien der Gattung Ocillatoria) nekrotische „Opferzellen“ (en. sacrificial cells), genannt Nekridien (oder Necridia), die durch ihr Absterben den Bruch ermöglichen. Sie unterbrechen dadurch die Kommunikation zwischen den beiden Teilen eines Filaments unterbrechen, so dass jeder Abschnitt seine Gleitrichtung ändern kann, was in der Folge zum Auseinanderbrechen führt.[32][4]
Die durchschnittliche Filamentlänge, die durch diesen Prozess erreicht wird, ist auch das Ergebnis von Wechselwirkungen zwischen Genen und Umwelt. Beispielsweise hängen das Wachstum und Position des Filaments von vertikalen Gradienten von Sauerstoff und Sulfid ab. Daher wird vorgeschlagen, dass gute Umweltbedingungen paradoxerweise den Zelltod der Nekridien verursachen, um den Fadenbruch und damit die Reproduktion zu fördern.[32]
Die Gattung Beggiatoa besteht hauptsächlich aus chemolithotrophen, sulfid-oxidierenden Bakterien. Die Palette der möglichen Stoffwechselwege ist jedoch sehr vielfältig und reicht von der Heterotrophie bis zur Chemolithoautotrophie. Aufgrund dieser großen Variabilität können sich die verschiedenen Arten und Stämme dieser Gattung stark voneinander unterscheiden.[27]
In der Gattung Beggiatoa gibt es sowohl autotrophe als auch heterotrophe Stoffwechselprozesse. Autotrophe Beggiatoa führen die CO2-Fixierung durch den Calvin-Zyklus und den Einsatz des Enzyms RuBisCO durch. Letzteres ist bei obligaten und fakultativen Autotrophen unterschiedlich stark reguliert. So kann RuBisCO im obligat autotrophen Stamm Beggiatoa sp. MS-81-1c[33] nicht unterdrückt werden, während es im fakultativ autotrophen Stamm Beggiatoa sp. MS-81-6[34] stark reguliert wird, um vom autotrophen zum heterotrophen Wachstum und umgekehrt zu wechseln. Neben den autotrophen Stämmen sind die meisten Süßwasser-Beggiatoa-Stämme heterotroph und benötigen organische Substrate für ihr Wachstum. Insbesondere können viele von ihnen als mixotroph angesehen werden, da sie heterotroph wachsen und organische Verbindungen oxidieren; aber auch Sulfid oder andere reduzierte Schwefelverbindungen als Elektronendonatoren nutzen können. Durch diese Strategie kann organischer Kohlenstoff für die Vermehrung der Biomasse gespeichert werde, eine autotrophe CO2-Fixierung ist dann nicht erforderlich. Mixotrophie wird als trophische Modalität für viele Süßwasserstämme zwar vermutet, wurde aber bisher nur in einem marinen Beggiatoa-Stamm, dem bezeichneten B. sp. MS-81-6, gefunden.[27][28] Auch ein Stoffwechselweg zur Nutzung von C1-Verbindungen (Moleküle mit einem einzigen Kohlenstoff-Atom) wurde im Stamm B. leptomitoformis D-402T, durch umfassende Analyse seiner Genomik, Physiologie und Biochemie/Molekularbiologie aufgedeckt.[17]
Die Beggiatoa-Bakterien zeigen eine große Vielseitigkeit bei der Verwertung von Stickstoffverbindungen. Stickstoff kann einerseits ein Wachstumsfaktor sein oder (wie beispielsweise als Nitrat) ein Elektronenakzeptor für die anaerobe Atmung. Heterotrophe Süßwasser-Beggiatoa-Arten assimilieren Stickstoff für ihr Wachstum. Zu den Stickstoffquellen gehören Nitrat, Nitrit, Ammoniak, Aminosäuren (wie Alanin , Asparagin und Asparaginsäure), Harnstoff und Thioharnstoff, je nach der Fähigkeit der einzelnen Arten und Stämme.[5][27]
Autotrophe vakuolierte Beggiatoa sind in der Lage, Nitrat in ihren Vakuolen in der 20.000-fachen Konzentration des umgebenden Meerwassers zu speichern und es unter anoxischen Bedingungen als terminalen Elektronenakzeptor zu nutzen. Bei diesem Prozess, der als Dissimilatorische Nitratreduktion zu Ammonium (en. dissimilatory nitrate reduction to ammonium, DNRA) bezeichnet wird, wird Nitrat zu Ammonium reduziert. Die Fähigkeit, Nitrat als Elektronenakzeptor zu nutzen, ermöglicht die Besiedlung von anoxischen Umgebungen wie mikrobiellen Matten und Sedimenten. Mehrere Beggiatoa-Arten sind in der Lage, Stickstoff mit Hilfe des Enzyms Nitrogenase zu fixieren, z. B. Beggiatoa alba.[5][27]
Eines der charakteristischen Merkmale der Gattung Beggiatoa ist die Produktion von intrazellulären Schwefeleinschlüssen, die aus der Oxidation reduzierter Schwefelquellen (z. B. Schwefelwasserstoff, H2S) resultieren. In autotrophen Beggiatoa-Vertretern ist Sulfid eine Energie- und Elektronenquelle für die Kohlenstofffixierung und das Wachstum. Die Oxidation von Sulfid kann aerob oder anaerob erfolgen, d. h. sie kann mit der Reduktion von Sauerstoff oder mit der Reduktion von Nitrat gekoppelt sein. Der durch die Oxidation von Sulfid erzeugte Schwefel wird in internen Globuli gespeichert und kann wieder verwendet werden, wenn die Sulfidkonzentration sinkt. Die vorübergehende Speicherung von elementarem Schwefel (S0) erhöht also die Anpassungsfähigkeit eines Organismus und seine Toleranz gegenüber Veränderungen der Sulfid- und Sauerstoffkonzentration.[5][27]
Aerobe Oxidation von Sulfiden: 2 H 2 S + O 2 ⟶ 2 S 0 + 2 H 2 O {displaystyle {ce {2H2S + O2 -> 2S^0 + 2H2O}}} 
Anaerobe Oxidation von Sulfiden: 4 H 2 S + NO 3 − + 2 H + ⟶ 4 S 0 + NH 4 + + 3 H 2 O {displaystyle {ce {4H2S + NO3^- + 2H+ -> 4S^0 + NH4^+ + 3H2O}}} 
Es gibt auch einige Fälle von Chemoorganotrophie. So betreibt beispielsweise der Stamm Beggiatoa sp. 35Flor[35] normalerweise eine aerobe mit der Oxidation von Sulfid gekoppelte Atmung; aber unter anoxischen Bedingungen wird eine andere Art der Atmung aktiviert: Die Energie wird dann chemoorganotroph aus der Oxidation von Polyhydroxyalkanoaten (PHA) gewonnen – das sind organische Verbindungen, die zuvor durch CO2-Fixierung während des chemolithotrophen Wachstums mit Hilfe von Sauerstoff und Sulfid synthetisiert wurden. In diesem Fall ist der Elektronenakzeptor der in der Zelle gespeicherte elementare Schwefel (S0), so dass das Endprodukt Schwefelwasserstoff ist.[23]
Anaerobe Atmung: PHA + S 0 ⟶ CO 2 + H 2 S {displaystyle {ce {PHA + S^0 -> CO2 + H2S}}} 
Der Stamm B. sp. 35Flor ist auch in der Lage, Wasserstoff als alternativen Elektronendonator zu Sulfid zu nutzen. Dieser Oxidationsprozess kann Energie für Wartungs- und Assimilationszwecke liefern und kann den gespeicherten Schwefel zu reduzieren, wenn er übermäßig wird. Er kann allerdings den Organismen kein Wachstum verschaffen.[24]
Wasserstoff-Oxidation: H 2 + S 0 ⟶ H 2 S {displaystyle {ce {H2 + S0 -> H2S}}} 
Der Stoffwechsel von Beggiatoa umfasst die Verwendung von Phosphor in Form von Polyphosphat. Die Regulierung dieses Stoffwechsels hängt von den Umweltbedingungen ab. Eine sauerstoffreiche Umgebung führt zu einer Anhäufung von Polyphosphat, während Anoxie (in Verbindung mit einer steigenden Sulfidkonzentration) einen Abbau von Polyphosphat und seine anschließende Freisetzung aus den Zellen bewirkt. Das freigesetzte Phosphat kann sich dann als Phosphorit-Mineral in den Sedimenten ablagern, aber auch im Wasser gelöst bleiben.[27]
Wie beobachtet bilden Beggiatoa-Fadenbakterien dichte Matten auf Sedimenten in einer Vielzahl von Umgebungen, wo sie als weißliche Schicht erscheinen. Sie leben an der oxischen/anoxischen Grenzfläche, wo sie sowohl von der Anwesenheit von Schwefelwasserstoff (H2S) als auch von Sauerstoff (O2) profitieren. Die chemolithoautotrophen Stämme von Beggiatoa werden auch als wichtige Primärproduzenten in dunklen Umgebungen angesehen.[5]
Die äußerst große Anpassungsfähigkeit der Stoffwechselvorgänge einzelner Beggiatoa-Vertreter in dieser Bakteriengattung ist eine evolutionäre Folge der außerordentlichen Variabilität der Umweltbedingungen, in denen sie leben. Beggiatoa ist fast benthisch, ihre Vertreter können einerseits in marinen Umgebungen (B. sp. MS-81-6 und B. sp MS-81-1c), andererseits auch in Süßwasserumgebungen (B. alba) vorkommen und benötigen nur Sulfid oder Thiosulfid (das Anion S=S– oder das Radikal S=S-, wiktionary: thiosulfide) als Elektronendonator und Oxidationsmittel. Sie sind in der Regel in Lebensräumen mit hohem Schwefelwasserstoffgehalt zu finden, z. B. in kalten Quellen, Schwefelquellen, mit Abwasser verunreinigtem Wasser, in Schlammschichten von Seen und in der Nähe hydrothermalen Schloten der Tiefsee. Beggiatoa findet man auch in der Rhizosphäre (Wurzelbereich) von Sumpfpflanzen,[36][13] im Boden, in Meeressedimenten und auch in Mangrovenlagunen, wo sie zum Lipid-Anteil der Sedimente beitragen.[37][38] Die Süßwasserarten haben typische Lebensräume in Schwefelquellen, Gräben, Pfützen, Feuchtgebieten, Seesedimenten und in Reisfeldern, wo sie in Verbindung mit den Wurzeln der Reispflanzen wachsen können. Die im Meerwasser lebenden Beggiatoa sind in Regionen zu finden, in denen ihre Energiequelle (Sulfid oder Thiosulfid) verfügbar ist. Sie können sowohl aus anorganischen als auch aus organischen Quellen gewonnen werden und sind in der Regel bei mikrooxischen Bedingungen, d. h. einer sehr niedrigen Sauerstoffkonzentration anzutreffen.[5] Außerdem findet man diese Gattung von Gammaprotobakterien auch in lokalisierten Bereichen anaerober Zersetzung, wie z. B. auf Kadavern im Wald und auf dem Meeresboden der Tiefsee.[5]
Vakuolisierte Beggiatoa (Beggiatoa mit sehr großen Vakuolen im Vergleich zum gesamten Zellvolumen) sind in küstennahen Auftriebsgebieten (z. B. an den Küsten Perus und Chiles), in hydrothermalen Schloten der Tiefsee und in Kaltwasserschloten (en. cold seeps) sehr häufig anzutreffen. In diesen Umgebungen können Flockungsmatten wachsen und große Flächen bedecken, ihre haarähnlichen Filamente können eine Höhe von 30 cm über dem Substrat erreichen. In hydrothermalen Schloten der Tiefsee und in Cold Seeps können Beggiatoa in Fäden wachsen, die einen Durchmesser von bis zu 200 mm (Mikrometern) haben, was sie zu den größten derzeit bekannten Prokaryoten macht. Vakuolisierte Beggiatoa kommen auch in hypoxischen Meeresböden (mit wenig Sauerstoff) vor, wo die Filamente in den Sedimenten in einer Tiefe von etwa 2 bis 4 cm Zentimetern leben anzutreffen sind. In diesen Fällen können die bakteriellen Filamente von Beggiatoa der häufigste Teil der mikrobiellen Biomasse in den Sedimenten darstellen.[5]
Beggiatoa können auch in Salzsümpfen, Salinen und geothermisch aktiven Unterwasserhöhlen vorkommen. Einige Studien zu diesen Lebensräumen wurden in den Unterwasserhöhlen des dolomitischen Kalksteins in Capo Palinuro, Provinz Salerno (Italien) durchgeführt. Hier gibt es hydrothermale sulfidische Quellen, und der mikrobielle Biofilm steht in Verbindung mit den Bewegung dieser hydrothermalen Flüssigkeiten infolge der Gezeiten. Die in den Höhlen gefundenen Filament-Matten bestanden größtenteils aus Bakterien, die Beggiatoa, der verwandten Gattung Thiothrix, sowie Flexibacter (Klasse Cytophagia der Bacteroidetes) ähneln. Diese Beggiatoa-ähnlichen Filamente sind morphologisch denen ähnlich, die an Felsen und am Byssus der Muscheln aus den Hydrothermalquellen von Lucky Strike auf dem Mittelatlantischen Rücken gefunden wurden.[5]
Häufig können die Mikroorganismen der Gattung Beggiatoa komplexe mikrobielle Matten bilden, in denen sie zusammen mit vielen anderen fadenförmigen Bakterien, z. B. Cyanobakterien, leben. Die Cyanobakterien besetzen in der Regel die Oberflächenschicht der Matte und produzieren tagsüber eine große Menge an Sauerstoff, der aus der Photosynthese stammt.[5] Umgekehrt wachsen die Beggiatoa-Bakterien entlang der oxischen/anoxischen Grenzfläche zwischen Sauerstoff und Sulfid, d. h. unterhalb der phototrophen Mikroorganismen, wo sie (aufgrund des von ihnen akkumulierten elementaren Schwefels) weiße Flecken erzeugen.[5]
Nebenstehend eine Konzeptionszeichnung der Biomasseverteilung, nachdem sich die Organismen an unterschiedlich vorgegebene Lichtbedingungen angepasst haben („photische Zonen“), gemessen bei einer Bestrahlungsstärke der einfallenden Photonen von 1000 μmol/m²s (Mikromol Photonenpro Quadratmeter und Sekunde).[39] Die Oberflächenschichten der mikrobiellen Matte werden von dichten Populationen beweglicher Cyanobakterien (Microcoleus sp. und anderer fadenförmiger Arten) und fadenförmigen sulfidoxidierenden Bakterien (Beggiatoa spp.) dominiert. Unterhalb der schmalen photischen Zone befindet sich eine – aufgrund von enthaltenem Eisen(II)-sulfid (FeS) dunkle – Sedimentschicht. Im Detail:[39]
Rechts: Während der Dunkelakklimatisierung wird die gesamte Matte jedoch anoxisch, so dass die Beggiatoa an die Oberfläche der Matte wandern und eine weiße Schicht über den Cyanobakterien ausbilden. Damit entgehen sie den hohen Konzentrationen an Schwefelwasserstoff (H2S) und bleiben an der Sauerstoff/Sulfid-Grenzfläche (O2-H2S-Grenze), während die Cyanobakterien in einer dichten Schicht darunter blieben.[40][39]
Links: Bei schwachem Licht werden die oberen Schichten oxidiert und treiben die O2-H2S-Grenzfläche und damit Beggiatoa tiefer in die Matte, während sich die beweglichen Cyanobakterien bei den schwachen (nicht hemmenden) Lichtverhältnissen in Richtung der Mattenoberfläche bewegen.[39]
Mitte:Bei starkem Licht wird ein größerer Teil des Biofilms oxidiert, die Beggiatoa bewegen sich weiter nach unten; die beweglichen Cyanobakterien in den oberen Schichten beginnen sich ebenfalls etwas nach unten zu bewegen, um die hohen (für sie inhibitorischen) Lichtwerte an der Oberfläche zu vermeiden.[39]
Manchmal werden Beggiatoa-Matten auch durch Kieselalgen und grüne Euglenoiden angereichert.[41] Im Ökosystem des Guaymas-Beckens wurden auch noch andere Protisten wie Wimpertierchen und Dinoflagellaten in Verbindung mit den Matten gefunden; diese verzehren wahrscheinlich eine große Menge der bakterieller Biomasse.[42]
Da die mikrobiellen Matten bis zu 3 cm breit werden können, stellen sie eine Nahrungsquelle für viele Weidetiere dar. Ein Beispiel für einen Fressfeind ist das Wimpertierchen Colpidium, dessen Auftreten sogar ein Indikator für Wasserverschmutzung darstellt (siehe Colpidium §Indikator für die Wasserverschmutzung). Diese trophische Assoziation wurde auch in Mangrovensystemen beobachtet, wo Beggiatoa einen Teil der Meeressedimente bedecken. Diese Bakterien bilden dort einen wichtigen Beitrag zur Ernährung der Meiofauna, insbesondere von Rädertierchen (Rotifera), Vielborstern (Polychaeta), Fadenwürmern (Nematoda) und einigen Gruppen von Plattwürmern (Plathelminthes), Schlauchwürmern (hier: Aschelminthes) und Kiefermündchen (Gnathostomulida).[43] Es wurde eine bemerkenswerte Beziehung zwischen Nematoden und Beggiatoa festgestellt: Tatsächlich scheinen Nematoden die Entwicklung von Beggiatoa-Matten zu begünstigen, indem sie das Eindringen von Sauerstoff und die Diffusion von Nährstoffen in die Matte fördern.[44] Ein weiteres Beispiel für einen Fressfeind von Beggiatoa ist der blinde Höhlenkrebs Tethysbaena ophelicola (Ranzenkrebse) in der israelischen Ajalon-Höhle.
Außerdem sind viele Aasreste (Kadaver) mit Matten aus Beggiatoa-ähnlichen Fadenbakterien bedeckt, die anaerobe, sulfatreduzierende Bakterien überlagern. Die Kadaver locken an sich viele Aasfresser (Nekrophagen) an; aber wenn sie die Matte aufbrechen, wird Schwefelwasserstoff freigesetzt und vertreibt die Aasfresser. Daher können Beggiatoa auch als ein „Schutz“ der Kadaver (en. carrion defence) vor den Aasfressern betrachtet werden.[45]
Mehrere Arten weißer Schwefelbakterien der Familie Beggiatoaceae (bzw. Thiotrichaceae) sind in der Lage, Nitrat (Nitrationen, NO3–), das aus flachen Küstensedimenten stammt und für den Stoffwechsel von grundlegender Bedeutung ist, zu akkumulieren, zu transportieren, sowie in ihren Fäden zu speichern. Die Reduktion von Nitrationen zu Ammonium setzt die Oxidation von Schwefelwasserstoff (H2S) voraus. Dieses Sulfid stammt außer bei geothermischen Quellen aus dem darunter liegenden anaeroben Sediment – in diesem Sediment findet eine dissimilatorische Sulfatreduktion statt,[41] die zur Bildung einer suboxischen Zone führt, die durch ein positives Redoxpotential und nur geringe Konzentrationen von freiem H2S gekennzeichnet ist. In der Meeresumwelt sid diese Arten wichtig, da sie eine grundlegende Rolle bei der Regulierung der H2S- und NO3–- Menge spielen.
Die Tatsache, dass Beggiatoa in der Lage ist, Phosphor als Polyphosphat zu akkumulieren und anschließend unter anoxischen Bedingungen als Phosphat in die Wassersäule freizusetzen, kann die Verfügbarkeit von Phosphor für Produzent (Ökologie)Primärproduzenten erhöhen. Einige Studien über den Phosphorkreislauf und die Freisetzung von Phosphor in Verbindung mit Beggiatoa wurden in der Ostsee durchgeführt. Diese Studien haben gezeigt, dass die Reduktion von Sulfid durch diese Bakterien die Geschwindigkeit der Eisensulfidbildung in den Sedimenten verringern und somit die Fähigkeit des Sediments, Phosphor zu binden, erhöhen kann.[27]
Da Beggiatoa in verschmutzten Meeresumgebungen vorkommen und gedeihen, können sie als Bioindikator angesehen werden;[47] genauso wie ihre Fressfeinde wie das Wimpertierchen Colpidium. Beggiatoa und andere verwandte Fadenbakterien können Absetzprobleme in kommunalen Kläranlagen wie in industriellen Abwasserkläranlagen (etwa bei der Konserven-Herstellung, Papierherstellung, Brauerei und Müllerei) verursachen, was zu dem Phänomen des „Bulking“ ([en]) führt. Geeignete Beggiatoa-Stämme sind auch in der Lage, Schwefelwasserstoff im Boden zu entgiften und spielen eine Rolle bei der Immobilisierung von Schwermetallen.[41][48]
Die erfolgreichsten Anreicherungen für Beggiatoa spp. wurden mit Hilfe einer flachen Schale oder eines Aquariums vorgenommen, in das einige Zentimeter Sand, verschiedene Mengen an Kalziumsulfat (CaSO4) und Dikaliumhydrogenphosphat (K2HPO4), eine Quelle komplexer organischer Polymere (Biopolymere) wie Algen, einige Zentimeter sulfidreicher Meeresschlamm und Meerwasser gegeben wurden. Die Anreicherung muss die richtige Sulfid-Sauerstoff-Grenzfläche aufweisen, was nur möglich ist, wenn Luft zugeführt wird, zum Beispiel durch einen langsamen, stetigen Fluss von frisch belüftetem Meerwasser.[5]
Eine andere Art der Anreicherung von Beggiatoa spp. basiert auf der Verwendung von extrahiertem getrocknetem Gras oder Heu in einem mineralischen Medium, da die komplexen Polymere wie Zellulosereste in diesem Material für die Sulfatreduktion sorgen. Dies liefert auch den Schwefelwasserstoff, der für die Anreicherung von Beggiatoa notwendig ist.[5]
Es gibt drei verschiedene Techniken, um isolierte Beggiatoa-Stämme in Reinkultur zu erhalten:
Dieses Verfahren zur Gewinnung eines isolierten heterotrophen Stammes erfordert eine Agarplatte mit verdünntem organischem Substrat, z. B. einer kleinen Menge Pepton. Dann müssen Büschel von Beggiatoa-Fäden aus der Umgebung gesammelt, mit steriler Waschlösung gewaschen und auf die Agarplatte gelegt werden (Inokulation). Auf diese Weise entstehen einige wachsende Fäden, die sich von dem zentralen Inokulum entfernen und als Inokulum für eine neue Agarplatte verwendet werden können. Für die Isolierung mariner Beggiatoa-Stämme (die autotrophes Wachstum zeigen) ist es für diese obligat mikroaereophile Organismen unerlässlich, mikrooxische Bedingungen (mit ein wenig Sauerstoff) zu schaffen und spezielle Agarplatten zu verwenden, die aus gefiltertem Meerwasser mit Natriumsulfid (Na2S) und Natriumacetat (NaC(O)CH3) hergestellt werden. Im Gegensatz dazu muss die Isolierung der Süßwasserstämme unter oxischen Bedingungen (Luftatmosphäre) erfolgen, wobei verschiedene Medien mit einer niedrigen Konzentration einer einzelnen organischen Verbindung wie Na2S, Acetat, oder Thiosulfat (S2O32−) verwendet werden.[5]
Flüssigmedien werden häufig zur Anreicherung, Titrierung (MPN-Auszählung, most probable number) und Massenkultivierung von Beggiatoa verwendet. Für eine erfolgreiche Kultivierung von heterotrophen oder mixotrophen Süßwasser-Beggiatoa müssen Flüssigmedien geringe Mengen an Kohlenstoffsubstrat enthalten, entweder Bodenextrakte oder Acetat. Der Referenzstamm der Typusart (B. alba str. B18LD) und verwandte Stämme werden im Allgemeinen in Medien kultiviert, die eine Salzbasis, Acetat als Kohlenstoffquelle und variable Hefeextrakt- und Sulfidzusätze enthalten.[49] Einige marine autotrophe Beggiatoa-Stämme werden auch auf definierten flüssigen mineralischen Medien mit Thiosulfat, CO2 unter mikro-oxischen Bedingungen – Belüftung mit 0,25 % O2 (v/v) in der Gasphase – kultiviert.[5]
Autotrophe Stämme, die aus einer Einzelfilamentisolierung auf Agar stammen, können leicht in Sulfidgradientenröhrchen (Röhrchen mit Konzentrationsgefälle ans Sulfid, vgl. auch Winogradsky-Säule) gehalten und vermehrt werden. Dabei werden sulfidreiche Agarpfropfen mit sulfidfreiem Weichagar überlagert. Die Röhrchen sind locker verschlossen, um den Austausch von Gasen oben im Kopfraum mit der Atmosphäre zu ermöglichen. Dadurch bilden sich zwei vertikal gegenüberliegende Schichten, von denen die eine Sulfid und die andere Sauerstoff enthält: Dies ermöglicht das Wachstum einer gut definierten Beggiatoa-Schicht an der sich ausbildenden Sulfid-Sauerstoff-Grenzfläche. Der Aufbau des Gradientenmediums erfordert unterschiedliche Mengen an J3-Medium – hergestellt aus Agar und Natron (NaHCO3) – das mit neutralisiertem Natriumsulfid (Na2S) ergänzt wird und in einem Schraubverschlussröhrchen untergebracht ist. Hier wird die Schwefelquelle durch den Sulfidfluss bereitgestellt. Eine weitere „Schicht“ besteht aus NaHCO3 ohne Sulfid oder Thiosulfat: Das gesamte Sulfid befindet sich unterhalb der Grenzfläche zwischen dem sulfidischen Agarpfropfen und dem übergelagerten sulfidfreien Agar, während sich im oberen Teil des Röhrchens eine weitere Schicht befindet, die das Sauerstoffreservoir darstellt. Aufgrund der Reaktion zwischen Sulfid und Sauerstoff beginnt sich ein Gefälle zu bilden: die Fäden vermehren sich schnell an der Sulfid-Sauerstoff-Grenzfläche und bilden eine markante Schicht oder „Platte“ mit einer Stärke von ca. 1 mm. Man sehen, dass diese Bakterien der Grenzfläche folgen und aufgrund der allmählichen Erschöpfung des Sulfid-Reservoirs langsam absteigen.[5]
Beggiatoa ist eine Gattung von Gammaproteobakterien, die zur Ordnung Thiotrichales (alias Beggiatoales) gehören. Diese Bakteriengattung war eine der ersten, die der russische Botaniker Sergei Winogradski (englisch auch Winogradsky) entdeckte. Während seiner Forschungen im botanischen Labor von Anton de Bary im Jahr 1887 stellte er fest, dass Beggiatoa Schwefelwasserstoff (H2S) als Energiequelle oxidiert und dabei intrazelluläre Schwefeltröpfchen bildet, wobei Sauerstoff der terminale Elektronenakzeptor ist und CO2 als Kohlenstoffquelle dient. Winogradski bezeichnete diese Form des Stoffwechsels als „Inorgoxidation“ (Oxidation anorganischer Verbindungen), heute Chemolithotrophie genannt. Die Beggiatoa-Arten leben in schwefelreichen Umgebungen auf Böden, sowohl im Süßwasser als auch im Meer, etwa in den hydrothermalen Schloten der Tiefsee und in verschmutzten Meeresumgebungen. Diese Entdeckung war die erste Entdeckung einer Lithotrophie. Diese farblosen Bakterien bilden fadenförmige Kolonien (Filamente, wie bei filamentösen Cyanobakterien auch Trichome genannt) und kann in einem Biofilm angeordnet sein, der mit bloßem Auge sichtbar ist und aus sehr langen weißen Fadenmatten besteht; die weiße Farbe ist dabei auf den gespeicherten Schwefel zurückzuführen. Die Beggiatoa-Bakterien sind dabei manchmal mit anderen Schwefelbakterien vergesellschaftet, z. B. mit der Gattung Thiothrix Winogradsky 1888 aus derselben Familie Thiotrichaceae Garrity et al. 2005.
Formell beschrieben wurden zwei Arten von Beggiatoa: die Typusart Beggiatoa alba und Beggiatoa leptomitoformis, wobei letztere erst 2017 veröffentlicht wurde. Diese Arten von Beggiatoa haben Zellen mit einem Durchmesser von bis zu 200 µm und gehören damit zu den größten Prokaryoten der Erde.
Beggiatoa is a genus of Gammaproteobacteria belonging to the order Thiotrichales, in the Pseudomonadota phylum. This genus was one of the first bacteria discovered by Ukrainian botanist Sergei Winogradsky. During his research in Anton de Bary's laboratory of botany in 1887, he found that Beggiatoa oxidized hydrogen sulfide (H2S) as an energy source, forming intracellular sulfur droplets, oxygen is the terminal electron acceptor and CO2 is used as a carbon source. Winogradsky named it in honor of the Italian doctor and botanist Francesco Secondo Beggiato (1806 - 1883), from Venice.[3] Winogradsky referred to this form of metabolism as "inorgoxidation" (oxidation of inorganic compounds), today called chemolithotrophy. These organisms live in sulfur-rich environments such as soil, both marine and freshwater, in the deep sea hydrothermal vents and in polluted marine environments. The finding represented the first discovery of lithotrophy.[4][5] Two species of Beggiatoa have been formally described: the type species Beggiatoa alba and Beggiatoa leptomitoformis, the latter of which was only published in 2017.[2][6] This colorless and filamentous bacterium, sometimes in association with other sulfur bacteria (for example the genus Thiothrix), can be arranged in biofilm visible to the naked eye formed by a very long white filamentous mat, the white color is due to the stored sulfur. Species of Beggiatoa have cells up to 200 µm in diameter and they are one of the largest prokaryotes on Earth.[7]
The genera Beggiatoa is a quite diverse group as it has representatives occupying several habitats and niches, both in fresh and salt water. In the past they have been confused as close relatives of Oscillatoria spp. (Cyanobacteria) for the morphology and motility characters,[8] but 5S rRNA analysis showed that members of Beggiatoa are phylogenetically distant from Cyanobacteria being members of the Gammaproteobacteria phylum.[9]
The capability to oxidize sulfide and store sulfur are the main features which separates Beggiatoa and closely related Thioploca as filamentous colorless sulfur bacteria from other filamentous bacteria (like cyanobacteria and the nonsulfur-oxidizing Cytophaga and Flexibacter)[10] Another defining feature is the ability to store nitrate inside the vacuoles of the wide marine species’ cells. 16S rRNA sequences base studies inferred that this characteristic is shared between members of a monophyletic clade nested in the Beggiatoa genera; this clade also includes members of Thioploca and Thiomargarita, both presenting only slight differences with Beggiatoas: whereas the former grows sharing a common slime sheath, the latter has not conserved filamentous growth and forms chains of rounded cells. Since the phylogenic history do not reflect the nomenclature, there's a need for a new denomination of genera and species.[11] The Neo-type strain is the B18LB and it settled the criteria for identification of the freshwater species Beggiatoa alba.[12]
According to NCBI database only two species of Beggiatoa spp. have been validly published: Beggiatoa alba, and Beggiatoa leptomitoformis.
Because of the lack of pure culture, little is known about the genetic of Beggiatoa. Beggiatoa alba show a GC content between 40 and 42.7 mol%, the presence of two or three similar plasmids and the genome size of Beggiatoa alba strain B18LD result to be about 3Megabase (Mb).[7] An important study about the genomic of Beggiatoa analyzed the sequences obtained from two single filaments of a vacuolated strain. Optical mapping showed that the genome size was about 7.4Mb and the sequence analyses detected pathways for sulfur oxidation, nitrate and oxygen respiration, and CO2 fixation, confirming the chemolithoautotrophic physiology of Beggiatoa. Furthermore, comparative genomics indicates horizontal gene transfer between Beggiatoa and Cyanobacteria of storage, metabolic, and gliding abilities.[13]

By the observation of their morphological characteristics, Beggiatoa spp. can be divided into three categories:
Obviously, this classification is purely ideal so some exceptions can exist. Narrow filaments are usually composed by cylindrical cells which length is about 1.5 to 8 times their thickness; wider filaments instead are disk-shaped with cell lengths from 0.10 to 0.90 times their cell width. In all of the cultured strains the terminal cells of the filaments appear rounded. Although they are Gram-negative bacteria, Beggiatoa show unusual cell-wall and membrane organization. Sometimes are present further membranes that cover the peptidoglycan layer and the number of this additional membranes is very variable. Their presence maybe is due to the harsh conditions in which some of these organisms live. Even the intracellular granules can be covered by extra-membranes structure. Beside the sulfur granules, the cells often show the presence of similarly stored granules of polyhydroxybutyrate and polyphosphate. Very common in large marine vacuolated Beggiatoa are hollow-structured filaments, composed by cells with a narrow cytoplasm surrounding a large central vacuole, exploited for nitrate storing.[7][14]
The filaments move by gliding and this movement is likely connected to string-like structures in the outer membrane and trans-peptidoglycan channels. Sometimes the filaments can also break through the formation of necridia cells in the middle of the filament. The motility of the filament is very important for the adaptability of the bacteria, because it allows to move on more suitable conditions for the cellular metabolism. The main drivers that guide the movement of Beggiatoa filaments are high oxygen and sulfide levels and light exposure, from which the filaments move away.[14]

Beggiatoa reproducing strategy is fragmentation. The growth of a colony leading to mat development is obtained through alternating filament elongation and breakage. Breakage can happen essentially in the middle of a stretched filament, at the tip of a filament loop or where a tip of a loop was once placed. The presence of sacrificial cells is fundamental as they interrupt the communication between two parts of one filament; in this way each section can change its gliding direction causing the split.
The average filament length achieved through this process is also result of gene-environment interactions as, for instance, the growth and position of the filament is function of vertical gradients of oxygen and sulfide. Therefore, it is proposed that good environmental conditions will paradoxically cause cell death in order to enhance filament breakage, thus reproduction.[15]
Beggiatoa group is mainly composed by chemolithotrophic, sulfide-oxidizing bacteria. However, the range of possible metabolic pathways is very diversified, varying from the heterotrophy to the chemolithoautotrophy. Because of this huge variability the diverse bacteria of this genus can differ greatly from each other.[14]
In Beggiatoa group are present both autotrophic and heterotrophic metabolisms. Autotrophic Beggiatoa carry out the CO2 fixation through the Calvin cycle and the employment of the RuBisCO enzyme. The latter shows different regulation levels in obligated and facultative autotrophs. For instance, in the obligately autotrophic strain MS-81-1c RuBisCO cannot be repressed, while in the facultatively autotrophic strain MS-81-6 it is tightly regulated to switch from autotrophic to heterotrophic growth and vice versa. Beside the autotrophic strains, most of the freshwater Beggiatoa strains are heterotrophic, requiring organic substrates for growth. Specifically, many of them can be considered mixotrophs, because they grow heterotrophically, oxidizing organic compounds, but they can also use sulfide or other reduced sulfur compounds as electron donors. By this strategy, the organic carbon skeletons are saved for the purpose of increasing biomass and the CO2 autotrophic fixation is not required. Mixotrophy has been suspected to be the trophic modality for many freshwater strains, but it has only been found in one marine strain of Beggiatoa, MS-81-6.[14] Also a metabolic pathway of C-1 compounds utilization has been revealed in Beggiatoa leptomitoformis strain D-402, through comprehensive analysis of its genomic, biochemistry, physiology and molecular biology.[16]
Beggiatoa group shows substantial versatility in utilizing nitrogen compounds. Nitrogen can be a source for growth or, in the case of nitrate, it can be an electron acceptor for anaerobic respiration. Heterotrophic freshwater Beggiatoa spp. assimilate nitrogen for growth. Nitrogen sources include nitrate, nitrite, ammonia, amino acids, urea, aspartate, asparagine, alanine and thiourea, depending on the capability of specific strains.
Autotrophic vacuolated Beggiatoa are able to store nitrate in their vacuoles 20.000 times the concentration of the surrounding sea water, and use it as terminal electron acceptor in anoxic conditions. This process, called Dissimilatory Nitrate Reduction to Ammonium (DNRA), reduces nitrate to ammonium. The capability of using nitrate as electron acceptor allows the colonization of anoxic environments, such as microbial mats and sediments. Several species are able to fix nitrogen using nitrogenase enzyme (e.g. Beggiatoa alba).[7][14]

One of the defining features of the genus Beggiatoa is the production of intracellular inclusions of sulfur resulting from the oxidation of reduced sulfur sources (e.g. hydrogen sulfide). In autotrophic Beggiatoa, sulfide is a source of energy and electrons for carbon fixation and growth. The oxidation of sulfide can be aerobic or anaerobic, in fact it can be coupled with the reduction of oxygen or with the reduction of nitrate. Sulfur produced by the oxidation of sulfide is stored into internal globules and can be used when the concentration of sulfide decreases. Thus, the temporarily storing of elemental sulfur (S0) increase the adaptability of an organism and its tolerance to changes in the concentrations of sulfide and oxygen.[7][14]
Sulfide aerobic oxidation: H 2 S + 0 , 5 O 2 ⟶ S 0 + H 2 O {displaystyle {ce {H2S + 0,5O2 -> S^0 + H2O}}} 
Sulfide anaerobic oxidation: 4 H 2 S + NO 3 − + 2 H + ⟶ 4 S 0 + NH 4 + + 3 H 2 O {displaystyle {ce {4H2S + NO3^- + 2H+ -> 4S^0 + NH4^+ + 3H2O}}}
There are some cases of chemoorganotrophy, too. For instance, the strain Beggiatoa sp. 35Flor usually do an aerobic respiration coupled with the oxidation of sulfide, but in anoxic condition a different type of respiration is activated. The energy is gained chemoorganotrophically from oxidation of PHA (polyhydroxyalkanoates), organic compounds previously synthesized through CO2 fixation during chemolithotrophic growth on oxygen and sulfide. In this case electron acceptor is the sulfur stored into the cell, so the final product is hydrogen sulfide.[17]
Anaerobic respiration: PHA + S 0 ⟶ CO 2 + H 2 S {displaystyle {ce {PHA + S^0 -> CO2 + H2S}}}
The strain Beggiatoa sp. 35Flor is able to use hydrogen as alternative electron donor to sulfide. This oxidation process can provide energy for maintenance and assimilatory purposes and is helpful to reduce stored sulfur when it becomes excessive, but it can't provide growth to the strain.[18]
Hydrogen oxidation: H 2 + S 0 ⟶ H 2 S {displaystyle {ce {H2 + S^0 -> H2S}}} 
Beggiatoa’s metabolism include the use of phosphorus in the polyphosphate form. The regulation of this metabolism relies on the environmental conditions. Oxygenated surroundings cause an accumulation of polyphosphate, while anoxia (coupled with an increasing concentration of sulfide) produces a breakdown of polyphosphate and its subsequent release from the cells. The released phosphate can then be deposited as phosphorite minerals in the sediments or stay dissolved in the water.[14]
Filaments have been observed to form dense mats on sediments in a very huge variety of environments. They appear as a whitish layer and since they are present and flourish in marine environments which have been subject to pollution, they can be considered as an indicator species.[19] Beggiatoa and other related filamentous bacteria can cause settling problems in sewage treatment plants, industrial waste lagoons in canning, paper pulping, brewing, milling, causing the phenomenon called "bulking". Beggiatoa are also able to detoxify hydrogen sulfide in soil and have a role in the immobilization of heavy metals.[20][21]
Beggiatoa live at the oxic/anoxic interface, where they benefits from the presence of both hydrogen sulfide and oxygen. The chemolithoautotrophic strains of Beggiatoa are also considered important primary producers in dark environments.[7]
The incredible number of adaptations and metabolisms of this genus of bacteria are consequences of the extraordinary environmental variability they can live in. Beggiatoa is almost benthic, it can be found in marine (Beggiatoa sp. MS-81-6 and MS-81-1c) or freshwater (Beggiatoa alba) environments and they only need sulfide or thiosulfide as electron donor and an oxidizer. They can usually be found in habitats that have high levels of hydrogen sulfide, these environments include cold seeps, sulfur springs, sewage contaminated water, mud layers of lakes, and near deep hydrothermal vents. Beggiatoa can also be found in the rhizosphere of swamp plants,[22][23] in soil, marine sediments and in the mangrove lagoon too (where they contribute to the lipid pool of the sediments[24]).[25] The freshwater species have typical habitats in sulfur springs, ditches, puddles, wetlands, lake sediments and in rice fields, where it can grow associated with the rice plants’ roots. The Beggiatoa that live in marine water can be found in regions where their source of energy (sulfide or thiosulfide) is available. It can be extracted from both inorganic or organic source and usually it is coupled with microoxic condition, therefore very low concentration of oxygen.[7] This genus of Gammaprotobacteria is also common in localizated area of anaerobic decomposition as whale carcass on the deep ocean seafloor.[7]
Vacuolated Beggiatoa can be very common in coastal upwelling regions (for example Peru and Chile coasts), deep sea hydrothermal vents and cold vents; in these environments the floc mats (hair-like breast) can grow up and cover large areas and reach the height of 30 cm. In deep sea hydrothermal vents and cold-seeps Beggiatoa can grow in filaments that can be up to 200 micrometres in diameter, which makes these ones the largest prokaryotes currently known. Vacuolated Beggiatoa can be found also in hypoxic seafloor, where the filaments can live inside the sediments at the depth of few cm (from 2 to 4 cm); in same cases the Beggiatoa bacterial filaments can be the most abundant part of the microbial biomass in the sediments.[7]
Beggiatoa can also be found in salt marshes, saline, and geothermally active underwater caves. Some studies on these environments have been carried out in the underwater caves of dolomitized limestone in Capo Palinuro, Salerno, (Italy). Here there are hydrothermal sulphidic springs and microbial biofilm is associated with the flow of hydrothermal fluids, whose activity is intermittent and starts during low tide. Mats found in the caves were composed by filaments resembling in most part Beggiatoa, Thiothrix and Flexibacter, and this Beggiatoa-like filaments were morphologically close to those found attached to rocks and the byssus of the mussels from Lucky Strike Hydrothermal vents on the Mid-Atlantic Ridge.[7]
Frequently, microorganisms of the genus Beggiatoa can form complex microbial mats, where they live in association with many other filamentous bacteria, such as cyanobacteria. The latter usually occupy the surface layer of the mat and during the day they produce a great amount of oxygen, derived from the photosynthetic activity. Conversely, Beggiatoa grow along an oxic/anoxic (oxygen/sulfide) interface, beneath the phototrophs, where they produce white patches.[7] However, during dark acclimation, the mat became anoxic, so the Beggiatoa moved to the mat surface, to avoid the high levels of H2S and remain at the oxygen/sulfide interface, while cyanobacteria remained in a dense layer below.[26] Sometimes Beggiatoa mats are enriched by the presence of diatoms and green euglenoids too,[20] but also protists as ciliates and dinoflagellates have been found associated with the mats at the Guaymas Basin hydrothermal vent ecosystem and they likely consume a large amount of bacterial biomass.[27]
As the microbial mats can reach 3 cm in width, they can represent a source of food for many grazers. This trophic connection has been observed in mangrove systems, where Beggiatoa cover part of marine sediments. It has been observed that these bacteria give an important contribution to meiofauna's diet, in particular rotifers, polychaetes, nematodes and some groups of platyhelminthes, aschelminths and gnathostomulids.[28] A remarkable relationship has been found between nematodes and Beggiatoa. In fact nematodes seem to favor development of Beggiatoa mats, through the increasing of oxygen penetration and nutrient diffusion into the mat.[29]
Furthermore, many carrion appear covered by mats of Beggiatoa-like filamentous bacteria overlying anaerobic sulfate-reducing bacteria. They attract many metazoans scavengers, but when they break the mat, it releases hydrogen sulphide that drive away the scavengers. Hence, Beggiatoa can also be considered a carrion defence from the scavengers.[30]
Several species of white sulfur bacteria in the family Beggiatoaceae are able to accumulate and transport NO3−, taken from shallow coastal sediments which is fundamental in metabolism, as well as accumulate it in filaments. The reduction of NO3− to ammonium implies the oxidation of H2S (except for geothermal vents, the sulphide derives from the underlying anaerobic sediment in which dissimilatory sulphate reduction occurs[20]): this reduction leads to the formation of suboxic zones characterized by positive redox potential and only trace concentrations of free H2S. In marine environment, the presence of these species is important because they have a fundamental role in regulation of the amount of H2S and NO3− :
Beggiatoa can also accumulate phosphorus as polyphosphate and it subsequently releases phosphate in anoxic conditions. This might increase the availability of phosphorus to primary producers if the phosphate is released from the sediment to the water column. Some studies about the phosphorus cycling and the release of phosphorus linked to Beggiatoa have been realized in Baltic Sea. These studies showed that the reduction of sulfide by these bacteria may decrease the rate of iron sulfide formation in the sediments, and thus increase the phosphorus retention capability of the sediment.[14]
The most successful enrichments for Beggiatoa spp. have been made using a shallow pan or aquarium in which it has been added few centimeters of sand, different amount of CaSO4 and K2HPO4, a source of complex organic polymers such as seaweed, several centimeters of sulfide-rich marine mud and seawater. The enrichment must contain the proper sulfide-oxygen interface that can be possible only if air is introduced, for example, by a slow steady flow of freshly aerated seawater. Another type of enrichment associated with Beggiatoa spp. is based on the use of extracted dried grass or hay in a mineral medium because the complex polymers such as cellulose residues in the material provide sulfate reduction. This also provides the hydrogen sulfide necessary to enrich for Beggiatoa.[7]
There are three different possible techniques to obtain isolated Beggiatoa strains in pure culture:
The procedure that allows to obtain isolated heterotrophic strain requires an agar plate containing dilute organic substrate such as small amount of peptone. Then, it's necessary to collect tufts of Beggiatoa filaments from the environment, to wash them using sterile washing solution and to place them on the agar plate. In this way there will be some growing filaments moving away from the central inoculum that can be used as inoculum for a new agar plate. For the isolation of marine Beggiatoa strains (that show autotrophic growth), since they are obligate microaerophiles it's essential to provide micro-oxic conditions and to use particular agar plates made by filtered seawater supplemented with sodium sulfide and sodium acetate. On the contrary, for the freshwater strains isolation it's necessary to perform under oxic conditions (air atmosphere) using a variety of media containing a low concentration of single organic compound such as acetate, Na2S or thiosulfate.[7]
Liquid media are often used for enrichment, MPN enumeration and bulk cultivation of Beggiatoa. In order to have a successful cultivation of heterotrophic or mixotrophic freshwater Beggiatoa, liquid media has to contain little amounts of carbon substrate, either soil extracts or acetate. The type species and strain (Beggiatoa alba str. B18LD) and related strains are generally grown in media that include a salt base, acetate as carbon source, and variable yeast extract and sulfide additions.[32] Some marine autotrophic Beggiatoa strains are also been cultured on defined liquid mineral medium with thiosulfate, CO2, micro-oxic conditions under aeration with 0.25% O2 (v/v) in the gas phase.[7]
Autotrophic strains coming from a single filament isolation on agar can easily be maintained and propagated in sulfide gradient tubes in which sulfide-rich agar plugs are overlaid with sulfide-free soft agar. Tubs are loosely closed in order to permit the exchange of headspace gasses with the atmosphere. As result, two opposite layers are formed, one that contains sulfide while the other one oxygen: this allows the growth of a well-defined Beggiatoa layer at the sulfide-oxygen interface. The gradient medium construction requires different amounts of J3 medium (made by agar and NaHCO3) supplemented with neutralized Na2S placed in a screw-capped tube. Here, the sulfur source is provided by the flux of sulfide. Another ‘’ layer ‘’ is made by NaHCO3 without sulfide or thiosulfate: all of the sulfide will be below the interface between the sulfidic agar plug and the sulfide-free overlay agar while there will be another layer in the top of the tube that represents the oxygen reservoir. It begins to form a gradient shape due to the reaction between sulfide and oxygen: as results, the filaments rapidly proliferate at the sulfide-oxygen interface, forming a marked layer or “plate,” of 1 mm but it's also possible to appreciate that these bacteria can track the interface and slowly descend owing to the gradual depletion of the sulfide reservoir.[7]
{{cite journal}}: Cite journal requires |journal= (help) Beggiatoa is a genus of Gammaproteobacteria belonging to the order Thiotrichales, in the Pseudomonadota phylum. This genus was one of the first bacteria discovered by Ukrainian botanist Sergei Winogradsky. During his research in Anton de Bary's laboratory of botany in 1887, he found that Beggiatoa oxidized hydrogen sulfide (H2S) as an energy source, forming intracellular sulfur droplets, oxygen is the terminal electron acceptor and CO2 is used as a carbon source. Winogradsky named it in honor of the Italian doctor and botanist Francesco Secondo Beggiato (1806 - 1883), from Venice. Winogradsky referred to this form of metabolism as "inorgoxidation" (oxidation of inorganic compounds), today called chemolithotrophy. These organisms live in sulfur-rich environments such as soil, both marine and freshwater, in the deep sea hydrothermal vents and in polluted marine environments. The finding represented the first discovery of lithotrophy. Two species of Beggiatoa have been formally described: the type species Beggiatoa alba and Beggiatoa leptomitoformis, the latter of which was only published in 2017. This colorless and filamentous bacterium, sometimes in association with other sulfur bacteria (for example the genus Thiothrix), can be arranged in biofilm visible to the naked eye formed by a very long white filamentous mat, the white color is due to the stored sulfur. Species of Beggiatoa have cells up to 200 µm in diameter and they are one of the largest prokaryotes on Earth.
Beggiatoa es un género de bacterias del orden de Thiotrichales. Está nombrado en honor del italiano médico y botánico F.S. Beggiato. Estos organismos viven en entornos ricos en azufre. Beggiatoa fue descubierta por el botánico ruso Sergei Winogradsky cuando trabajó en el laboratorio de Botánica de Anton de Bary en 1887. Winogradsky encontró que Beggiatoa oxida sulfuro de hidrógeno (H2S) como fuente de energía, formando gránulos de azufre elemental intracelulares. Winogradsky refirió a esta forma de metabolismo como litotrofia[2][3] (oxidación de compuestos inorgánicos). Así, este hallazgo representa el primer descubrimiento de litotrofia. Se han descrito dos especies de este género, Beggiatoa alba[4] y Beggiatoa leptomitoformis.[5]
Beggiatoa puede ser encontrado en ambientes marinos o de agua dulce, normalmente en hábitats con niveles altos de sulfuro de hidrógeno, incluyendo incluyen aguas residuales contaminadas, capas de barro de lagos y zonas con fumarolas hidrotermales profundas. Beggiatoa también puede ser encontrado en el rizosfera de plantas de ciénagas.[6][7]
Son células incoloras con forma de disco o cilindro que forman filamentos largos con un diámetro de célula que puede medir entre 12 y 160 micrómetros (subespecies diferentes). Una vacuola central masiva se utiliza para acumulación de nitrato, presumiblemente para ser un aceptor de electrones en la oxidación anaerobia del sulfuro. Los filamentos están rodeados por lodo y puede mover por deslizamiento (sliding).
Beggiatoa puede crecer de forma quimioorganotrofa oxidando compuestos orgánicos a dióxido de carbono en presencia de oxígeno, a pesar de que altas concentraciones de oxígeno pueden ser un factor limitante. Los compuestos orgánicos son también la fuente de carbono para la biosíntesis. Alguna especie pueden oxidar el sulfuro hidrógeno a azufre elemental como una fuente suplementaria de energía (litoheterotrofía facultativa). El azufre producido se almacena intracelularmente.[8]
Beggiatoa es un género de bacterias del orden de Thiotrichales. Está nombrado en honor del italiano médico y botánico F.S. Beggiato. Estos organismos viven en entornos ricos en azufre. Beggiatoa fue descubierta por el botánico ruso Sergei Winogradsky cuando trabajó en el laboratorio de Botánica de Anton de Bary en 1887. Winogradsky encontró que Beggiatoa oxida sulfuro de hidrógeno (H2S) como fuente de energía, formando gránulos de azufre elemental intracelulares. Winogradsky refirió a esta forma de metabolismo como litotrofia (oxidación de compuestos inorgánicos). Así, este hallazgo representa el primer descubrimiento de litotrofia. Se han descrito dos especies de este género, Beggiatoa alba y Beggiatoa leptomitoformis.
Beggiatoa é un xénero de bacterias da orde Thiotrichales. Recibe o seu nome polo médico e botánico italiano F. S. Beggiato. Estes microorganismos viven en ambientes ricos en xofre. O botánico ruso Sergei Winogradsky, durante as súas investigacións no laboratorio de botánica de Anton de Bary en 1887, descubriu que Beggiatoa oxidaba sulfuro de hidróxeno (H2S) como fonte de enerxía, formando pinguiñas de xofre intracelular. Winogradsky denominou esta forma de metabolismo inorgoxidación[1] (oxidación de compostos inorgánicos). Este descubrimento representou o primeiro caso descrito de litotrofia.[2][3]
Beggiatoa pode atoparse en ambientes mariños e de auga doce. Poden xeralmente atoparse en hábitats que teñen altos niveis de sulfuro de hidróxeno. Estes ambientes inclúen emanacións frías hidrotermais mariñas, fontes sulfurosas, auga contaminada por vertidos residuais, capas de lodo, e nas proximidades de chemineas hidrotermais profundas. Beggiatoa pode tamén encontrarse na rizosfera de plantas de pantanos.[4][5]
As células de Beggiatoa son incoloras e discoidais ou cilíndricas, e dispóñense en longos filamenos cun diámetro celular entre 12 e 160 micrómetros (segundo as distintas subespecies). Utilizan un vacúolo central masivo para acumular nitrato, que seguramente usan como aceptor de electróns na oxidación anaerobia do sulfuro.[4] Os filamentros están rodeados por un limo e poden moverse escorregando.
Beggiatoa pode crecer de modo quimioorganótro ou heterótrofo oxidando compostos orgánicos a dióxido de carbono en presenza de oxíxeno, aínda que as altas concentracións de oxíxeno poden ser un factor limitante. Os compostos orgánicos son tamén a fonte de carbono para a biosíntese. Algunhas especies poden oxidar o sulfuro de hidróxeno a xofre elemental como unha fonte suplementaria de enerxía (facultativamente lito-heterótrofa). O xofre producido é almacenado intracelularmente.[6]
Algunhas especies teñen a capacidade de crecemento quimiolitoautótrofo por medio da oxidación do sulfuro para obter enerxía e usando o dióxido de carbono como fonte de carbono para as biosínteses. Neste metabolismo o nitrato almacenado internamente é o aceptor de electróns e é reducido a amoníaco.
As especies de Beggiatoa autótrofas mariñas poden oxidar o xofre intracelular a sulfato.[6]
Un mecanismo que se dá frecuentemente cando falta o oxíxeno é a redución do xofre elemental. O xofre redúcese a sulfuro a costa do carbono almacenado ou ao engadir gas hidróxeno. Isto pode ser unha estratexia de supervivencia para pasar os períodos sen oxíxeno.[6]
Os filamentos poden formar densos tapetes sobre os sedimentos dos ambientes de estuarios, plataformas continentais, emanacións de augas mariñas, e chemineas hidrotermais. Teñen o aspecto de capas abrancazadas e, ao estaren presentes en abundancia en ambientes mariños que foron polucionados, poden considerarse unha especie indicadora.
Beggiatoa e outras bacterias filamentosas poden causar problemas de depósitos indesexados en plantas de tratamento de augas residuais, lagoas de almacenamento de residuos industriais en plantas conserveiras, de pasta de papel, destilerías ou fundicións, causando un fenómeno chamado "bulking". As Beggiatoa poden tamén detoxificar solos contaminados por sulfuro de hidróxeno.
O xénero Beggiatoa está estreitamente emparentado con Thioploca.[4]
Beggiatoa é un xénero de bacterias da orde Thiotrichales. Recibe o seu nome polo médico e botánico italiano F. S. Beggiato. Estes microorganismos viven en ambientes ricos en xofre. O botánico ruso Sergei Winogradsky, durante as súas investigacións no laboratorio de botánica de Anton de Bary en 1887, descubriu que Beggiatoa oxidaba sulfuro de hidróxeno (H2S) como fonte de enerxía, formando pinguiñas de xofre intracelular. Winogradsky denominou esta forma de metabolismo inorgoxidación (oxidación de compostos inorgánicos). Este descubrimento representou o primeiro caso descrito de litotrofia.
Il genere Beggiatoa comprende batteri in grado di vivere in ambienti ricchi di zolfo. Prende il nome dal medico e botanico italiano Francesco Secondo Beggiato.
Beggioatoa är ett släkte bland de vita svavelbakterierna. I äldre tid hänfördes de av vissa till de blågröna algerna.
Organismen består av en ogrenad, jämntjock, fritt rörlig, färglös tråd, innehållande svavelkorn, och förekommer i såväl sött som salt vatten på svavelvätehaltig gyttjebotten eller i svavelkällor. Beggiatoa alba kan när den förekommer talrikt täcka bottnen som ett vitt överdrag över stora områden.
Beggioatoa är ett släkte bland de vita svavelbakterierna. I äldre tid hänfördes de av vissa till de blågröna algerna.
Organismen består av en ogrenad, jämntjock, fritt rörlig, färglös tråd, innehållande svavelkorn, och förekommer i såväl sött som salt vatten på svavelvätehaltig gyttjebotten eller i svavelkällor. Beggiatoa alba kan när den förekommer talrikt täcka bottnen som ett vitt överdrag över stora områden.
Beggiatoa (лат.) — род гамма-протеобактерий из порядка Thiotrichales[en]. Представители рода населяют местообитания, богатые серой. Именно у бактерий этого рода в 1887 году русский исследователь С. Н. Виноградский открыл литотрофию[1][2].
Род Beggiatoa получил своё название в честь Франческо Секондо Беджато (итал. Francesco Secondo Beggiato; 1806—1883), итальянского врача и ботаника из Виченцы, автора труда Delle Terme Euganea[3].
Представители рода Beggiatoa распространены по всему миру и населяют пресные и морские воды. Часто они живут в местообитаниях, обогащённых сероводородом. К таким местообитаниям относятся холодные источники[en], серные источники, сточные воды, озёрный ил, а также глубоководные геотермальные источники. Они также могут обнаруживаться в ризосфере болотных растений[4][5]. В очень солёных водах филаменты Beggiatoa совместно с бактериями других видов могут образовывать бактериальные маты[6].
Beggiatoa — многоклеточные (нитчатые) бесцветные бактерии, отдельные клетки имеют цилиндрическую (пресноводные и морские штаммы) или дисковидную (крупные морские штаммы) форму, а их диаметр составляет от 12 до 160 мкм (филаменты пресноводных штаммов существенно тоньше). Длина филамента может быть от нескольких мм до 10 см. Это одна из крупнейших известных бактерий. Имеется крупная центральная вакуоль, использующаяся для запасания нитрата. Белый цвет клеток обусловлен накоплением в клетке элементарной серы[7]. Кроме включений серы, в цитоплазме клеток также обнаруживаются включения поли-β-гидроксибутирата и пирофосфата. Бактерии подвижны и демонстрируют хемотаксис в ответ на градиент различных природных соединений. В отличие от близкородственного рода Thioploca, бактерии рода Beggiatoa не выделяют общего полисахаридного чехла, окутывающего пучок филаментов: каждая нить образует вокруг себя слизистый матрикс, состоящий из полисахаридов и белковых конъюгатов[6][4].
Хотя проверенные штаммы Beggiatoa окрашиваются по Граму отрицательно, строение их клеточной стенки весьма необычно для грамотрицательных бактерий. Она состоит по крайней мере из двух слоёв различного состава: один слой состоит в основном из пептидогликана и липополисахаридов, а второй имеет филаментозную структуру. Филаментозный слой играет ключевую роль в скользящем движении нитей Beggiatoa, которые могут скользить со скоростью 1—3 мкм/с. Для скольжения им необходим контакт с твёрдым субстратом[6].
Beggiatoa может расти хемоорганогетеротрофно, окисляя органические соединения до СО2 в присутствии кислорода, хотя высокие концентрации кислорода могут служить лимитирующим фактором роста бактерий. Органические соединения также служат источником углерода для биосинтетических процессов. Некоторые представители могут окислять Н2S до элементарной серы, используя этот процесс как вспомогательный источник энергии (факультативные литогетеротрофы)[8]. Окисление Н2S до серы и образование внутриклеточных серных включений у Beggiatoa было открыто русским учёным С. Н. Виноградским в 1887 году, и это был первый описанный случай литотрофии[1][2].
Некоторые представители рода могут расти хемолитоавтотрофно, используя окисление Н2S до серы как источник энергии, а углекислый газ как источник углерода для биосинтетических процессов. Акцептором в этом случае является нитрат, запасаемый в центральной вакуоли и восстанавливающийся до аммония. Многие автотрофные представители рода Beggiatoa могут окислять внутриклеточную серу до сульфата[8]. Некоторые штаммы могут в аэробных условиях окислять водород, а в анаэробных условиях водород используют для серного дыхания. Окисление водорода может быть не только энергетическим подспорьем, но также препятствовать чрезмерному отложению серы в цитоплазме[9].
Ближайшими родственниками Beggiatoa являются бактерии рода Thioploca[4].
По данным List of Prokaryotic names with Standing in Nomenclature[en], в состав рода Beggiatoa включают 2 вида[3]:
Геном штамма Beggiatoa leptomitiformis D-402T был полностью секвенирован[11].
Beggiatoa (лат.) — род гамма-протеобактерий из порядка Thiotrichales[en]. Представители рода населяют местообитания, богатые серой. Именно у бактерий этого рода в 1887 году русский исследователь С. Н. Виноградский открыл литотрофию.
ベギアトア属(学名:Beggiatoa)はチオスリックス科に属するグラム陰性の非芽胞形成偏性好気性桿菌。滑走運動をする硫黄細菌の一種で化学合成独立栄養生物であるが炭素固定に必要な酵素が不足しているため有機化合物を利用する混合栄養の株が多い。電子供与体として硫化水素のほか、チオ硫酸イオンを利用できる。名称はイタリアの物理学者、フランチェスコ・ベッギアートに因む。GC含量は37から51。
水中に生息し、pH6から8の範囲で増殖し硫化水素の存在下でよく見られる。体内に単体硫黄の顆粒を持つ。また、目視で確認できるほどの大きさを持つ。排水処理におけるバルキングの原因菌としても知られる。
ベギアトア属(学名:Beggiatoa)はチオスリックス科に属するグラム陰性の非芽胞形成偏性好気性桿菌。滑走運動をする硫黄細菌の一種で化学合成独立栄養生物であるが炭素固定に必要な酵素が不足しているため有機化合物を利用する混合栄養の株が多い。電子供与体として硫化水素のほか、チオ硫酸イオンを利用できる。名称はイタリアの物理学者、フランチェスコ・ベッギアートに因む。GC含量は37から51。
水中に生息し、pH6から8の範囲で増殖し硫化水素の存在下でよく見られる。体内に単体硫黄の顆粒を持つ。また、目視で確認できるほどの大きさを持つ。排水処理におけるバルキングの原因菌としても知られる。













{kind=link}