الأسماء في صفحات التنقل

Els paramecis (Paramecium) són uns organismes microscòpics ciliats amb forma de sola de sabata, habituals en aigües dolces estancades amb abundant matèria orgànica, com bassals i estancs.[1] Són probablement els eucariotes unicel·lulars més coneguts i segur que els protozous ciliats més estudiats per la ciència. La mida de les espècies sol rondar els 0,05 mil·límetres pel que calen microscopis per observar-los amb nitidesa.
Estan mancats de flagels, però els cilis són molt abundants i recobreixen tota la superfície. Els correspon proporcionar moviment a l'organisme sent utilitzats com rems.[1] La membrana externa absorbeix i expulsa regularment l'aigua de l'exterior amb la fi de controlar l'osmoregulació, procés dirigit per dos vacúols contràctils.
De la seva ultraestructura destaca el citostoma, una espècie d'invaginació situada al llarg del parameci que empra per a la captura d'aliment, conformat per partícules orgàniques flotants i microorganismes menors. El citostoma condueix a una citofaringe abans que l'aliment passi a l'interior d'aquest protozou. Altres orgànuls de fàcil observació són el típic nucli eucariota, situat junt a un "micronucli" en el centre del parameci, i les vaquoles digestives, que digereixen constantment l'aliment capturat. Les desfetes s'expulsen per exocitosi, mitjançant vacúols de secreció que s'originen a partir de les digestives.
S'alimenta de bacteris i altres microorganismes. Té una boca en forma de ranura.[1]
Com molts altres microorganismes, els paramecis es reprodueixen asexualment per bipartició.
Es reconeixen 14 espècies dintre del gènere Paramecium:
Els paramecis (Paramecium) són uns organismes microscòpics ciliats amb forma de sola de sabata, habituals en aigües dolces estancades amb abundant matèria orgànica, com bassals i estancs. Són probablement els eucariotes unicel·lulars més coneguts i segur que els protozous ciliats més estudiats per la ciència. La mida de les espècies sol rondar els 0,05 mil·límetres pel que calen microscopis per observar-los amb nitidesa.
Estan mancats de flagels, però els cilis són molt abundants i recobreixen tota la superfície. Els correspon proporcionar moviment a l'organisme sent utilitzats com rems. La membrana externa absorbeix i expulsa regularment l'aigua de l'exterior amb la fi de controlar l'osmoregulació, procés dirigit per dos vacúols contràctils.
De la seva ultraestructura destaca el citostoma, una espècie d'invaginació situada al llarg del parameci que empra per a la captura d'aliment, conformat per partícules orgàniques flotants i microorganismes menors. El citostoma condueix a una citofaringe abans que l'aliment passi a l'interior d'aquest protozou. Altres orgànuls de fàcil observació són el típic nucli eucariota, situat junt a un "micronucli" en el centre del parameci, i les vaquoles digestives, que digereixen constantment l'aliment capturat. Les desfetes s'expulsen per exocitosi, mitjançant vacúols de secreció que s'originen a partir de les digestives.
S'alimenta de bacteris i altres microorganismes. Té una boca en forma de ranura.
Com molts altres microorganismes, els paramecis es reprodueixen asexualment per bipartició.
Trepka (Paramecium) je rod obrvených vodních prvoků z kmene nálevníci, třídy chudoblanní.[1] Mnoho znaků sdílí s ostatními zástupci svého kmene, ale má mnoho jiných, unikátních vlastností. Charakteristický je tvar boty[2], který dal tomuto prvokovi i české rodové jméno.
Z organel jsou patrná buněčná ústa (cytostom), kde vznikají potravní vakuoly, dále například stažitelná vakuola. Z metabolického hlediska jsou to heterotrofové, fagocytují bakterie; někdy však mohou obsahovat zelené endosymbionty. Rozmnožují se pohlavně i nepohlavně (pohlavní proces je tzv. konjugace). Trepka velká se živí např. planktonem, bakteriemi nebo velice drobnými živočichy.
Rozeznáváme mnoho druhů rodu Paramecium. Známé jsou:
Trepka se rozmnožuje většinou nepohlavně. Jedinec vyroste na dospělém jedinci a po čase se oddělí. Trepky mají i pohlavní rozmnožování. Nazývá se konjugace. Jedinci splynou buněčnými ústy, generativní jádro prodělá redukční dělení, vegetativní se dvakrát rozpadá. Následuje vznik čtyř jader, tři z nich zanikají a čtvrté se haploidně rozdělí na dvě. Jádra v buňkách splynou v synkarion a konjuganti se rozestoupí. Následují tři za sebou jdoucí jaderná dělení. Výsledkem konjugace je vznik osmi trepek.
Trepka (Paramecium) je rod obrvených vodních prvoků z kmene nálevníci, třídy chudoblanní. Mnoho znaků sdílí s ostatními zástupci svého kmene, ale má mnoho jiných, unikátních vlastností. Charakteristický je tvar boty, který dal tomuto prvokovi i české rodové jméno.
Z organel jsou patrná buněčná ústa (cytostom), kde vznikají potravní vakuoly, dále například stažitelná vakuola. Z metabolického hlediska jsou to heterotrofové, fagocytují bakterie; někdy však mohou obsahovat zelené endosymbionty. Rozmnožují se pohlavně i nepohlavně (pohlavní proces je tzv. konjugace). Trepka velká se živí např. planktonem, bakteriemi nebo velice drobnými živočichy.
Die Pantoffeltierchen (Paramecium) sind eine Gattung von rundum bewimperten Protisten. Das Merkmal „Zilien“ („Wimpern“) haben die Pantoffeltierchen mit einigen anderen Einzellern gemeinsam, die man traditionell zu den Wimpertierchen (Ciliata, Ciliophora) zählt. Als erste Protisten ernannte die Deutsche Gesellschaft für Protozoologie die Vertreter der Gattung Paramecium im Jahr 2007 zu den Einzellern des Jahres.
Pantoffeltierchen leben vorwiegend im Süßwasser, einige Arten wie Paramecium woodruffi sind aber euryhalin und kommen auch im Brackwasser von Flussmündungen, selten sogar im Meer, vor.[1] Die meisten Vertreter trifft man in Gewässern wie z. B. Tümpeln, Teichen, Seen, Flüssen, aber auch in Wasserpfützen an. Somit sind sie ein wichtiger Bestandteil des Ökosystems Süßwasser. Pantoffeltierchen sind häufig vorkommende Ciliaten. Die Gattung Paramecium bildet im Gegensatz zu vielen anderen Einzellern keine Cysten als Überdauerungsstadien aus.[2]
Pantoffeltierchen sind frei lebende, sich aktiv bewegende Einzeller mit einer Länge zwischen 50 und 300 Mikrometern. In Aufsicht ist ihre Körperform langgestreckt oval („zigarrenförmig“) bis rundlich, wobei das Vorderende mehr oder weniger asymmetrisch schief abgeschnitten (also sohlen- oder „pantoffelförmig“) wirkt. Im Wasser vermögen sie sich frei schwimmend schnell zu bewegen, wobei sie sich fast immer nach links spiralförmig um ihre Achse drehen. Von oben nach unten (dorsoventral) ist der Zellleib abgeplattet. Auf der Unterseite (Ventralseite) sitzt ein grubenförmiges Mundfeld. Diese grubenförmige Einsenkung zieht sich – in etwa halber Körperlänge – asymmetrisch vom linken Vorderende bis zur Körpermitte. Am Ende geht die Einsenkung in den Mundtrichter (Vestibulum) über, an dessen spitzem Ende eine Mundöffnung (der Zellmund) liegt. Vom Zellmund aus zieht sich eine röhrenartige Buccalhöhle, die in Aufsicht meist s-förmig erscheint und mit spitzem Ende blind endet, nach innen. Die Anordnung und Morphologie der Cilien in der Buccalhöhle ist wichtig für die genaue Bestimmung der Arten. Die Zelloberfläche wird durch eine steife, aber dabei elastische Hülle, Pellicula genannt, verstärkt. Diese ist auf der Oberseite in langgestreckt sechseckige (hexagonale) Felder gegliedert. Aus jedem der Felder entspringt eine, selten auch zwei Zilien („Wimpern“), die reihig auf der gesamten Oberfläche sitzen. Bei einigen Arten sind sie am Hinterende büschelartig verlängert. Unter der Pellikula liegt ein dünner, Rinde (Cortex) genannter Bereich des Zellplasmas, in dem die Trichocysten sitzen (vgl. unten im Kap. Verteidigung). Das darunter liegende Endoplasma wirkt dünnflüssiger und enthält zahlreiche Organellen, oft auch Kristalle, die sich mit der Plasmaströmung im Zellkörper mitbewegen. Normalerweise sind zwei kontraktile Vakuolen vorhanden. Diese pulsieren alternierend (abwechselnd) und geben ihr Inneres durch eine Pore nach außen ab. Im typischen Fall sind zwei Arten von Zellkernen vorhanden: ein einzelner, großer Makronukleus von elliptischer bis nierenförmiger Gestalt, ein wenig seitlich in der Körpermitte gelegen, und eine je nach Art unterschiedliche Anzahl viel kleinerer Mikronuklei, die meist in unmittelbarer Nachbarschaft liegen.[3]
Die größten Arten zählt man zu den „Riesen“ unter den Einzellern, da sie schon mit bloßem Auge als kleine, weiße Pünktchen in einem Wassertropfen zu erkennen sind.
Pantoffeltierchen fallen unter dem Mikroskop durch schnelles Umherschwimmen auf. Sie sind außen von vielen (etwa 10.000) Wimpern (Zilien) umgeben, die der Fortbewegung dienen. Durch das rhythmische Krümmen und Wiederaufrichten der Wimpern ziehen sich Schlagwellen über den Körper des Pantoffeltierchens. Durch die spiralförmige Anordnung dieser Wimpern wird das Pantoffeltierchen um seine Längsachse gedreht, wodurch die körperbedingte seitliche Bahnabweichung nicht zur Kreisbewegung, sondern zur Schraubenbahn wird. Es hat für einen Einzeller eine recht große Geschwindigkeit von 1 bis 1,4 mm/s. Hindernisse oder Engpässe passiert das Pantoffeltierchen mit Leichtigkeit, da es sich aufgrund einer elastischen Zellmembran (genauer: Pellicula) mühelos an ihnen vorbei- oder hindurchbewegen kann. Drehungen in jede Richtung sind möglich, bei Schreckreaktionen sogar durch Umkehr des Cilienschlages ein plötzliches Rückwärtsschwimmen.
Das Pantoffeltierchen nimmt seine Beute und Nahrung durch seinen „chemischen“ Sinn (Chemorezeptoren) und durch Tastreize (andere molekulare Rezeptoren) wahr. Es ernährt sich vorwiegend von Bakterien, die durch Wimpernschläge zum Mundfeld befördert werden. Die Wimpern wirken also auch bei der Nahrungsaufnahme mit, indem sie Nahrungspartikel heranstrudeln. Mit Hilfe der Mundfeldbewimperung gelangen die Bakterien über das Mundfeld zum Zellmund, wo sie anschließend im Zellschlund in eine Nahrungsvakuole importiert werden. Dieser Vorgang heißt Endocytose.
Zu dieser Nahrungsvakuole führen zahlreiche bandartige Strukturen, mithilfe derer viele Membranbläschen herantransportiert werden, die die Nahrungsvakuole vergrößern. Hat sie eine bestimmte Größe erreicht, schnürt sie sich ins Zellinnere ab. Überschüssiges Wasser wird der Nahrungsvakuole entzogen.
Zunächst gelangen Acidosomen in die Nahrungsvakuole und senken den pH-Wert auf 1,2 ab. Über Lysosomen gelangen Verdauungsenzyme ins Bläschen, das von nun an Verdauungsvakuole heißt. Während die Verdauung abläuft und die Bakterien zersetzt werden, wird die Verdauungsvakuole auf einer ovalförmigen Bahn durch die Zelle transportiert. Dies nennt man Cyclose.
Die verwertbaren Nahrungsstoffe werden resorbiert und die unverdaulichen Substanzen über die Cytopyge (den so genannten Zellafter) ausgeschieden. Damit dies geschehen kann, muss das Verdauungsbläschen als Kontaktvakuole am Zellafter Kontakt mit der Zellmembran aufnehmen. Dieser Vorgang heißt Exozytose. Da es nahezu ständig Nahrung in sich hineinstrudelt, kann ein Pantoffeltierchen innerhalb weniger Stunden seine Körpermasse verdoppeln.
In die Zelle eingedrungenes überschüssiges Wasser wird mit Hilfe von sternförmig angeordneten Zuführungskanälen in die Sammelblasen von zwei pulsierenden Bläschen (kontraktilen Vakuolen) geleitet und aus diesen über einen Exkretionsporus aktiv ausgeschieden.
Pantoffeltierchen vermehren sich normalerweise ungeschlechtlich durch Querteilung in zwei Tochterzellen. Das Pantoffeltierchen zieht sich in die Länge und das Mundfeld teilt sich. Es bildet sich jeweils eine weitere pulsierende Vakuole aus. Der Mikronukleus und der Makronukleus verdoppeln sich. Das Pantoffeltierchen schnürt nun noch den Zellleib so durch, dass jedes der beiden auf diese Weise neu entstehenden Individuen einen Kleinkern, einen Großkern, zwei pulsierende Vakuolen und ein Mundfeld enthält. Das Pantoffeltierchen teilt sich unter günstigen Bedingungen bis zu siebenmal pro Tag.[4]
Gelegentlich kommt es auch zu geschlechtlichen Vorgängen, die man Konjugation nennt und bei denen die Paramecien mit anderen Individuen der gleichen Art Erbinformationen austauschen. Zwei Pantoffeltierchen legen sich dazu an den Mundfeldern aneinander. Die Zellmembranen verschmelzen in diesem Bereich, die Wimpern verschwinden. Die Großkerne lösen sich allmählich auf. Die Kleinkerne teilen sich durch Reduktionsteilung in vier haploide Tochterkerne. Von diesen vier sterben drei ab, und einer teilt sich zu zwei haploiden Kernen. Jeweils ein Tochterkern wandert in das andere Pantoffeltierchen, um mit dem dort verbliebenen Tochterkern zu einem neuen diploiden Kern zu verschmelzen. In jedem Individuum teilt sich der durch Verschmelzung entstandene neue Kern in zwei Tochterkerne. Der eine ist der neue Kleinkern, der andere entwickelt sich (unter mehrfacher Verdoppelung und vollständiger Neuorganisation der Chromosomen) zu einem Großkern. Nun trennen sich die Konjugationspartner wieder, Wimpern und Mundfelder werden im fehlenden Abschnitt ergänzt.
Die Konjugation wird unter anderem durch jahreszeitliche Änderungen oder sich verschlechternde Umweltbedingungen stimuliert.
Das Pantoffeltierchen reagiert sehr stark auf Reize der Umgebung (Berührung, Temperatur, chemische Reize, Belichtung). In geschlossenen Glasröhren schwimmen sie immer in Richtung Wasseroberfläche, obwohl sie schwerer als Wasser sind. Sie reagieren auf Schwerkraft (Gravitaxis). Pantoffeltierchen nehmen chemische und thermische Reize nur mit dem vorderen Teil des Körpers wahr. Trifft ein Pantoffeltierchen auf ein Hindernis, schwimmt es durch Umkehrung des Wimpernschlages ein Stück zurück und vollführt eine leichte Drehung. Dann schwimmt es wieder nach vorn. Trifft es wieder auf das Hindernis, so probiert es das Pantoffeltierchen solange mit dieser Methode, bis es an dem Hindernis vorbeikommt. Das Pantoffeltierchen arbeitet koordiniert, da nicht nur die betroffene Stelle, sondern das ganze Lebewesen die Reaktion ausführt.
Zu den Feinden der Pantoffeltierchen gehören Amöben und Einzeller der Gattung Didinium. Letztere gehören ebenfalls zu den Ciliaten. Didinium ergreift mit Hilfe einer nasenartigen Vorstülpung das Pantoffeltierchen und verleibt es sich anschließend ein. Amöben und Sonnentierchen umhüllen das Pantoffeltierchen mit Scheinfüßchen und verdauen es anschließend in einer Nahrungsvakuole.
Gegen Angreifer versucht sich das Pantoffeltierchen mit Hilfe der Trichocysten zu wehren. Es handelt sich dabei um stäbchenförmige Gebilde enthaltende Haarbläschen, deren Spitzen kalzifiziert sind, die direkt unterhalb der Zellmembran liegen und bei Gefahr lange, klebrige Proteinfäden (Eiweißfäden) ausschleudern. Sobald ein Angreifer ein Pantoffeltierchen berührt, verlängern sich diese Gebilde explosionsartig und schießen die Proteinfäden ins Wasser. Ein ganzes Büschel der Proteinfäden kann dem Pantoffeltierchen helfen, sich Feinde vom Leib zu halten. In den abgeschossenen Proteinfäden können sich manche Fressfeinde verfangen und letztendlich auch absterben. Ausgestoßene Trichocysten werden durch neue ersetzt, die in Vesikeln im Cytoplasma entstehen.
Pantoffeltierchen werden als Indikator für die Gewässerverschmutzung genutzt.[5] Außerdem werden sie auch als Fischfutter verwendet.[6]
Wie viele Ciliaten sind Pantoffeltierchen kerndimorph, d. h. der Zellkern kommt in zwei verschiedenen Formen vor. Ein großer Zellkern, Makronukleus genannt, ist somatisch aktiv, in ihm werden Gene abgelesen und transkribiert. Ein oder mehrere kleine Zellkerne oder Mikronuklei sind in dieser Zeit in Ruhestadien. Ihre Gene werden nicht transkribiert. Bei der normalen, ungeschlechtlichen Zellteilung werden Makro- und Mikronukleus wie üblich separat, über eine Mitose, verdoppelt und auf die Tochterzellen verteilt. Bei der geschlechtlichen Fortpflanzung (einer Meiose), entweder über ein Konjugation zwischen zwei Zellen oder über Autogamie, wird der Makronukleus nicht verdoppelt, sondern zerstört. Anschließend wird aus einem Mikronukleus ein neuer Makronukleus gebildet.
Der genetische Gehalt in Makro- und Mikronukleus ist identisch, d. h. in ihnen sind im Prinzip dieselben Gene vorhanden. Allerdings wird das Genom bei der Bildung des Makronukleus vollkommen umgebaut. Alle Chromosomen werden mehr als achthundert Mal verdoppelt. Anschließend werden aus zahlreichen kurzen Sequenzstücken daraus funktionale Gene zusammengesetzt. Repetitive DNA und Transposonen werden ausgeschnitten. Durch den vollständigen Umbau ist der Makronukleus anschließend homozygot.
Da alle Kopien des Makronukleus bei einer geschlechtlichen Fortpflanzung verloren gehen, werden letztlich nur Mutationen im Mikronukleus dauerhaft vererbt. Dadurch besitzen Pantoffeltierchen eine äußerst geringe Mutationsrate.[7] Vielleicht erleichtert durch den eigenartigen Fortpflanzungsmodus wurden aber im Genom von Paramecium (untersucht wurde die Art Paramecium tetraurelia aus dem aurelia-Artkomplex) mit fast 40.000 eine ungewöhnlich hohe Anzahl von Genen gefunden. Dieses Pantoffeltierchen besitzt damit, überschlägig, etwa die doppelte Anzahl an Genen wie der Mensch, Homo sapiens. Die ungewöhnlich hohe Genanzahl wird auf mehrere Runden (mindestens drei) von Verdoppelung des gesamten Genoms zurückgeführt, bei der zahlreiche paraloge Gene entstehen.[8]
Die Gattung Paramecium gehört zu den Peniculia[9], einer Gruppe der Wimpertierchen, die je nach Autoren im Rang einer Unterordnung, einer Ordnung oder einer Unterklasse eingeordnet wird. Innerhalb der Gruppe steht sie nach genetischen Daten relativ isoliert mit hoher Distanz zu anderen, sie wird deshalb formal als einzige Gattung in die damit monotypische Familie Parameciidae gestellt, die meisten Bearbeiter verzichten aber ganz auf die Angabe einer Familie. Mögliche Schwestergruppe könnte die überwiegend marine Gattung Frontonia sein.[10][11]
Die Gattung wird nach morphologischen und genetischen Daten in vier[12], nach neueren Ergebnissen sogar fünf[13] Untergattungen geteilt. Die Abgrenzung und Anzahl der Arten anzugeben ist schwierig. Es werden etwas weniger als 20 Morphospezies unterschieden, die sich nach anatomischen Merkmalen unter dem Mikroskop unterscheiden lassen. Die Arten können teilweise anhand der generellen Körpergestalt[12], von langgestreckt „zigarrenförmig“ bis „pantoffelförmig“ mit in Aufsicht rundlicher Gestalt mit abgerundet schief abgeschnitten wirkendem Vorderende, gruppenweise unterschieden werden. Zwei Arten sind durch endosymbiontische Algen grün gefärbt. Eine genaue Bestimmung[14] beruht auf der Gestalt und Anordnung der Cilien, der Zahl der Mikronuklei und verschiedener Vakuloen. Innerhalb der Morphospezies werden aber zusätzlich Kryptospezies unterschieden, das sind Arten, die morphologisch gleich (oder fast gleich) aussehen, sich aber anhand von genetischen, biologischen und ökologischen Daten unterscheiden lassen. Bekannt ist insbesondere der „Paramecium aurelia“-Artenkomplex.[14] Was sich morphologisch als eine Morphospezies darstellt, konnte nach dem Paarungsverhalten in fünfzehn „Stämme“ oder „Linien“ geschieden werden, bei denen jeweils zwei Paarungstypen (o, nach englisch odd, ungleich und e, nach englisch even, gleich) existieren. Jeder dieser Stämme akzeptiert bei der Konjugation (im Wesentlichen) nur Individuen desselben Stamms, wobei sich ausschließlich o und e miteinander paaren – dadurch ist es im Labor möglich, durch entsprechende Auslese im Kulturmedium die Konjugation ganz zu unterdrücken. Die Stämme sind später von ihrem Entdecker als Arten beschrieben worden, wobei er sie im Namen einfach (lateinisch) durchnummerierte: Paramecium primaurelia, Paramecium biaurelia, Paramecium triaurelia usw. Da diese in der Reproduktion gegeneinander isoliert sind, ist ein wesentliches Kriterium für die Abgrenzung einer Art, trotz der fehlenden morphologischen Merkmale, erfüllt. Genetisch[15] konnte die Eigenständigkeit der kryptischen Arten des aurelia-Artenkomplexes im Wesentlichen bestätigt werden. Unerwarteterweise existierten aber einige Stämme, die nach der genetischen Verwandtschaft zu einer bestimmten Linie gehörten, sich aber mit Angehörigen eines „falschen“ Stammes paarten; hier wird ein späterer Wechsel durch eine nachträgliche Mutation angenommen. Auch andere Morphospezies wie Paramecium multimicronucleatum erwiesen sich als aus kryptischen Arten zusammengesetzt,[16] sind aber schlechter erforscht.
In der folgenden Aufstellung[17] sind nur die Morphospezies enthalten.
Untergattung Paramecium.
Untergattung Chloroparamecium
Untergattung Helianter
Untergattung Cypriostomum
Untergattung Viridoparamecium
Die Pantoffeltierchen (Paramecium) sind eine Gattung von rundum bewimperten Protisten. Das Merkmal „Zilien“ („Wimpern“) haben die Pantoffeltierchen mit einigen anderen Einzellern gemeinsam, die man traditionell zu den Wimpertierchen (Ciliata, Ciliophora) zählt. Als erste Protisten ernannte die Deutsche Gesellschaft für Protozoologie die Vertreter der Gattung Paramecium im Jahr 2007 zu den Einzellern des Jahres.
Parameci është një protozoarë (parashtazorë).
Është perfaqesuesi kryesor i qerpikoreve (ciliate ).
Leviz me ane te qerpikeve te shumte qe dalin nga trupi i tij.
Qerpiket sherbejne për te levizur dhe për te shtyre ushqimin drejt një hapesire te vogel ne forme hinke.
Ne fundin e saj formohet një fshikez ku futen ushqimet.
Fshikeza futet brenda trupit dhe ushqimet treten brenda saj.
Te tille protiste jetojne ne mjedise te ndotura dhe nuk shkaktojne sëmundje dhe shumohet me ndarje.
Paramecijum (lat.: Paramecium) je jedan od najpoznatijih rodova trepljara. Zbog svog oblika dobio je i naziv "papučica". Paramecijum je otkrio 1675. godine Antoni van Leeuwenhoek pod svojim jednostavnim mikroskopom.
Papučica je obavijena opnom koja se zove pelikula. Na pelikuli su brojne treplje kojima se papučica pokreće i uzima hranu. Pojedine treplje mogu biti malo jače i nepokretne. Pretpostavlja se da su to osjetila za mehaničke i hemijske nadražaje. Disanje se vrši cijelom površinom tijela. Ima dvije kontraktilne vakuole (služe za izbacivanje viška vode) i dva jedra - veliko i malo. Ona se razlikuju i po veličini, i po obliku.
Na pelikuli se vidi udubljenje koje nazivamo usno predvorje (vestibulum) koje završava citostom pomoću kojeg papučica prima hranljive materije. Čelijsko ždrijelo - citofarink se nastavlja na citostom i na njegovom se kraju stvaraju hranljivi mjehurići. Oni se spajaju s lizosomima koji razgrađuju hranu, a neprobavljeni ostaci se izbacuju na određenom mjestu na pelikuli koje zovemo čni izmetni otvor ili citopig. Papučica, slično kao i amebe, diše difuzno preko površine tijela. Količinu vode u organizmu papučice reguli še pomoću para stežljivih mjehurića (kontraktilnih vakuola). Svi trepljari imaju barem dvia jedra: malo i veliko. Razmnožavaju se binarnom diobom.
Kada naiđu nepovoljni uslovi, paramecijum obrazuje cistu oko sebe. Dok je unutar ciste, on se ne hrani i ne razmnožava. Kada prođu nepovoljni uslovi, on se oslobađa ciste i nastavlja normalan život.
Paramecium bursaria, vrsta sa simbiotskim algama
Paramecium putrinum
Dioba paramecijuma
Paramecium caudatum pri konjugaciji
Paramecium ya iku salah sawijining protista wujud kéwan. Paramecium duwé ukuran watara 50–350 cm. Paramecium nduwé ukuran kang luwih gedhé saka amoeba, dhapuré kaya pérangan ngisor sepatu. Awaké ditutupi déning rambut-rambut cilik kang digunaake kanggo obah. Rambut-rambut cilik mau karan silia. Paramecium obah nganggo siliane. [1]
Ciri-ciri saka paramecium ing antarané:
Paramecium ya iku salah sawijining protista wujud kéwan. Paramecium duwé ukuran watara 50–350 cm. Paramecium nduwé ukuran kang luwih gedhé saka amoeba, dhapuré kaya pérangan ngisor sepatu. Awaké ditutupi déning rambut-rambut cilik kang digunaake kanggo obah. Rambut-rambut cilik mau karan silia. Paramecium obah nganggo siliane.
Ciri-ciri saka paramecium ing antarané:
Nduwéni celah cangkem kang dipirantèni déning anus sèl. Nduwéni loro inti sel, ya iku makronukleus (piranti reproduksi aseksual) lan mikronukleus (piranti reproduksi sèksual). Ing pérangan dinding sel ndan rambut getar (silia) kanggo piranti gerak. Reproduksi sèksual kanthi cara konjugasi lan aseksual kanthi cara nyigar awak. Urip ing banyu anta kang ameh ngasal zat nonorganik.Paramecijum (lat.: Paramecium) je jedan od najpoznatijih rodova trepljara. Zbog svog oblika dobio je i naziv "papučica". Paramecijum je otkrio 1675. godine Antoni van Leeuwenhoek pod svojim jednostavnim mikroskopom.
Papučica je obavijena opnom koja se zove pelikula. Na pelikuli su brojne treplje kojima se papučica pokreće i uzima hranu. Pojedine treplje mogu biti malo jače i nepokretne. Pretpostavlja se da su to osjetila za mehaničke i hemijske nadražaje. Disanje se vrši cijelom površinom tijela. Ima dvije kontraktilne vakuole (služe za izbacivanje viška vode) i dva jedra - veliko i malo. Ona se razlikuju i po veličini, i po obliku.
Na pelikuli se vidi udubljenje koje nazivamo usno predvorje (vestibulum) koje završava citostom pomoću kojeg papučica prima hranljive materije. Čelijsko ždrijelo - citofarink se nastavlja na citostom i na njegovom se kraju stvaraju hranljivi mjehurići. Oni se spajaju s lizosomima koji razgrađuju hranu, a neprobavljeni ostaci se izbacuju na određenom mjestu na pelikuli koje zovemo čni izmetni otvor ili citopig. Papučica, slično kao i amebe, diše difuzno preko površine tijela. Količinu vode u organizmu papučice reguli še pomoću para stežljivih mjehurića (kontraktilnih vakuola). Svi trepljari imaju barem dvia jedra: malo i veliko. Razmnožavaju se binarnom diobom.
Kada naiđu nepovoljni uslovi, paramecijum obrazuje cistu oko sebe. Dok je unutar ciste, on se ne hrani i ne razmnožava. Kada prođu nepovoljni uslovi, on se oslobađa ciste i nastavlja normalan život.
பேரமீசியம் (இலங்கை வழக்கு: பரமீசியம் - paramecium) என்பது ஓரணுவுயிர் தொகுதியில் சாறு வாழ் உயிர்கள் வகுப்பைச் சார்ந்ததாகும். சேற்றில் தோன்றி வாழும் இவ்வகை உயிர்கள் (Infusoria) என அழைக்கப்படுகின்றன. இது மிதியடி வடிவச் சிற்றுயிர் ஆகும். பெரும்பாலான நீர் நிலைகளில் காணப்படும் இது பல மீன்களுக்கு உணவாகி அவற்றை வாழ வைக்கிறது.
பேரமீசியம் திட்டவட்டமான வடிவமைப்பைக் கொண்டது. குதியற்ற மிதியடியின் வடிவத்தைக் கொண்டுள்ள இது 0.25 மி.மீ வரை நீளமுடையது. இதனை வெறுங்கண்ணால் கூடப் பார்க்க முடியும். மினுமினுப்பான எண்ணற்ற நுண்மயிர்கள் இதன் உடலின் மேல் வளர்ந்துள்ளன. இதன் உடலின் முன்பாகம் குறுகியும் பின் பாகம் அகன்றும் காணப்படும். முன்பக்க நுனி அகன்றும் பின்பக்க நுனி கூர்மையாகவும் இருக்கும். பின்பக்க நுனியில் வால் போன்ற நீண்ட நுண்மயிர்கள் தோற்றமளிக்கும்.பின்னால் பெருகிய பல்லுயிரணு உயிர்களின் வால் அமைப்பிற்கான அடிப்படை ஓரணு உயிரான பேரமீசியத்தில் கணப்படுகிறது என்பது குறிப்பிடத்தக்கதாகும்.
பேரமீசியம் ஓரணு உயிர் என்ற போதும் பெரியதும் சிறியதுமான இரண்டு உட்கருக்களைக் கொண்டுள்ளது. பேரமீசியத்தின் உடல் உயிரணு ஊனீர்ப் படிவு என்னும் போர்வையால் மூடப்பட்டு உள்ளது. இப்போர்வையே சவ்வுறை எனப்படுகிறது. இந்தச் சவ்வுறையில் நுண்மயிர்கள், பிற உயிர்களிடமிருந்து தற்காத்துக் கொள்ள உதவும் குட்டை நுண்கோல்கள் முதலியவை உள்ளன.
பேரமீசியம் தன் உடல் முழுவதாலும் சுவாசிகிறது. காற்றிலுள ஆக்ஸிஜனை உட்கொண்டு கரியமில வாயுவை வெளியிடுகிறது. இதன் உடலில் பச்சையம் இல்லாததால் இது கரியமில வாயுவை உட்கொள்ள முடியாது.
இடைவிடாது நீரில் இயங்கிக் கொண்டிருக்கும் உயிர் பேரமீசியம் ஆகும். இதன் உடலைச் சுற்றியுள்ள நுண்மயிர்கள் துடுப்புகளைப் போல அசைந்து இயக்கத்திற்கு உதவுகின்றன். உணவைத் தேடிச் செல்லும் பேரமீசியம் தன் உடலின் நீண்ட அச்சைச் சுற்றிச் சுழன்றபடி நுண்மயிர்த் துடுப்புகளை அசைத்து அசைத்து நீரில் முன்னேறிச் சென்று தன் இயக்கத்தை நிகழ்த்துகிறது.
பேரமீசியங்கள் நீர்நிலைகளில் கரைந்துள்ள புல்நுண்ணுயிர்கள் எனப்படும் பாக்டீரியங்களைப் புசிக்கின்றன. பாக்டீரியங்களைப் புசிப்பதன் மூலம் இவை நீர்நிலைகளைச் சுத்தப்படுத்துகின்றன. பேரமீசியங்கள் தாமும் பல மீன்களுக்கு இரையாகின்றன.
பேரமீசியத்திற்கு வாய் இருக்கிறது. வாய்க்கு மேலே வாயருகுக் குழி ஒன்று உள்ளது. வாயருகுக் குழியில், நுண்மயிர்களை அசைப்பதன் மூலம் பல வகை உணவுத்துணுக்குகள் உட்செலுத்தப்படுகின்றன. வாயருகுக் குழியிலிருந்து உணவுப் பொருளானது, வாய்க்குச் செல்கிறது. வாயிலிருந்து உணவுக் குழலுக்குள் சென்று திரட்டப்பட்டு, கவள உருண்டை வடிவில், உணவு உயிரணு ஊனீரில் செலுத்தப்படுகிறது. பேரமீசியத்தின் உடலில் செரிப்புக் குமிழி உருவாகி அதனுள் உணவு சிறிது சிறிதாக செரிக்கபடுகிறது. செரிக்காத உணவுப் பொருள்கள் உடலின் பின்புறத்தே உள்ள மலத்துளை வழியே வெளியேற்றப்படுகின்றன.
செரிக்கப்படாத உணவுப் பொருள்கலை வெளியேற்ற மலத்துளை இருப்பதைத் தவிர வேறு வெளியேற்றக் குமிழிகளும் பேரமீசியத்தில் அமைந்துள்ளது ஒரு சிறப்பம்சமாகும். பேரமீசியத்தின் உயிரணு ஊனீரில் இரு வெள்ளிக் குமிழிகள் உள்ளன. இவை அடிக்கடி சுருங்கக்கூடியவை. இவற்றைச் சுற்றி விண்மீன் போன்றுல்ல ஆரைச் சீர் வடிகால்கள் உள்லன. உயிரணு ஊனீரில் சேரும் தேவையற்ற நீரும், நீர்ப்பொருள்களும் இந்த வடிகால்களில் நிரம்பி, பின் சுருங்குவதன் மூலம் வெளியேற்றப்படுகின்றன. இரண்டு வெள்ளிக் குமிழிகளும் ஒன்றன்பின் ஒன்றாகச் சுருங்கி, உடலுக்குத் தேவையில்லாத பொருள்களை வெளியேகொட்டுகின்ற அமைப்பு பேரமீசியத்தின் உடலில் காணப்படுகிறது.
பேரமீசியங்களில் இனப்பெருக்கம் இரண்டு வகைகளில் நிகழ்கிறது.
நன்கு வளர்ந்த பேரமீசியம் சாதகமான சூழ்நிலையில் பல்குகிறது. அப்போது அதன் உள்லேயுள்ள இரண்டு உட்கருக்களும் பாதியாகப் பிளவுறுகின்றன. கருக்கள் பிளவுறும் நிலையில் பேரமீசியத்தின் நடு உடலில் சுருக்கம் ஏற்பட்டு ஏறக்குறைய சமமான இரண்டு பகுதிகளாகத் தனித்தனியே ஓர் உயிரணுவைக் கொண்டவாறு பிரிந்து இனப்பெருக்கம் நிகழ்கிறது.
இரண்டு பேரமீசியங்கள் ஒன்றோடு ஒன்று பொருந்துவதன் மூலம் கலவி முறையில் இனப்பெருக்கம் நிகழ்கிறது. பேரமீசியங்கள் தங்கள் வாயருகுக் குழிகளை ஒன்றோடொன்று பொருத்திக் கொள்ளும் போது பெரிய உட்கருக்கள் கரைந்து சிறிய உட்கருக்கள் பல தடவை பிளவுபடுகின்றன. ஒன்றின் உட்கருப்பகுதிகள் இன்னொன்றின் உடலுக்குள் புகுகின்றன. இந்தக் கலவிக்குப்பின் குறிப்பிட்ட அவ்விரு பேரமீசியங்களில் புதிய உட்கருக்கள் பெரியதும் சிறியதுமாக உருவாகி இனப்பெருக்கத்திற்கு வித்திடுகின்றன.
பேரமீசியங்களை ஒரு பாத்திரத்தில் போட்டு, அப்பாத்திரத்தின் ஒரு பகுதியை மட்டும் சூடேற்றினால், 24 செ. முதல் 28° செ வெப்ப நிலையில் எல்லா பேரமீசியங்களும் ஒன்று திரண்டு நிற்கும். 30° செ -36 °செ -ல் பேரமீசியங்கள் அந்தப் பகுதியிலிருந்து விலகி குளிர்ச்சியான பகுதி நோக்கி விரையும். இதனால் 10°செ- 28° செ வரையுள்ள வெப்ப நிலையில் மட்டுமே பேரமீசியங்கள் உயிர் வாழ வல்லவை என்பதும் வெப்பத் தூண்டலுக்கேற்பத் துலங்கும் பண்புடையவை என்றும் தெரிகிறது. மேலும், பேரமீசியங்களின் நடுவே ஓர்உப்புத் துண்டைப் போட்டால் அவை விலகிப் போய்விடுகின்றன. ஆனால் ஒரு பாக்டீரியப் படலத்துணுக்கைப் போட்டால் அவை சூழ்ந்து மொய்த்து உண்ணுகின்றன. இதன் மூலம் பேரமீசியங்கள் புறத்தூண்டலுக்கேற்பச் செயல்படும் தன்மை கொண்டவை என்று தெரியலாம்.
ஆணைவாரி ஆனந்தன். 'பல்துறை அறிவியல்' மணியம் பதிப்பகம் வெளியீடு. 1989.
http://en.wikipedia.org/wiki/Paramecium
http://microbewiki.kenyon.edu/index.php/Paramecium
http://101science.com/paramecium.htm
http://www.bio.umass.edu/biology/conn.river/parameci.html
பேரமீசியம் (இலங்கை வழக்கு: பரமீசியம் - paramecium) என்பது ஓரணுவுயிர் தொகுதியில் சாறு வாழ் உயிர்கள் வகுப்பைச் சார்ந்ததாகும். சேற்றில் தோன்றி வாழும் இவ்வகை உயிர்கள் (Infusoria) என அழைக்கப்படுகின்றன. இது மிதியடி வடிவச் சிற்றுயிர் ஆகும். பெரும்பாலான நீர் நிலைகளில் காணப்படும் இது பல மீன்களுக்கு உணவாகி அவற்றை வாழ வைக்கிறது.
Paramecium (/ˌpærəˈmiːʃ(i)əm/ PARR-ə-MEE-sh(ee-)əm, /-siəm/ -see-əm; also spelled Paramoecium, plural Paramecia)[1] is a genus of eukaryotic, unicellular ciliates, commonly studied as a model organism of the ciliate group. Paramecia are widespread in freshwater, brackish, and marine environments and are often abundant in stagnant basins and ponds. Because some species are readily cultivated and easily induced to conjugate and divide, they have been widely used in classrooms and laboratories to study biological processes. The usefulness of Paramecium as a model organism has caused one ciliate researcher to characterize it as the "white rat" of the phylum Ciliophora.[2]


Paramecia were among the first ciliates to be observed by microscopists, in the late 17th century. They were probably known to the Dutch pioneer of protozoology, Antonie van Leeuwenhoek, and were clearly described by his contemporary Christiaan Huygens in a letter from 1678.[3] The earliest known illustration of a Paramecium species was published anonymously in Philosophical Transactions of the Royal Society in 1703.[4]
In 1718, the French mathematics teacher and microscopist Louis Joblot published a description and illustration of a microscopic poisson (fish), which he discovered in an infusion of oak bark in water. Joblot gave this creature the name "Chausson", or "slipper", and the phrase "slipper animalcule" remained in use as a colloquial epithet for Paramecium, throughout the 18th and 19th centuries.[5]
The name "Paramecium" – constructed from the Greek παραμήκης (paramēkēs, "oblong") – was coined in 1752 by the English microscopist John Hill, who applied the name generally to "Animalcules which have no visible limbs or tails, and are of an irregularly oblong figure."[6] In 1773, O. F. Müller, the first researcher to place the genus within the Linnaean system of taxonomy, adopted the name Paramecium, but changed the spelling to Paramœcium. C. G. Ehrenberg, in a major study of the infusoria published in 1838, restored Hill's original spelling for the name, and most researchers have followed his lead.[7]

Species of Paramecium range in size from 50 to 330 micrometres (0.0020 to 0.0130 in) in length. Cells are typically ovoid, elongate, or foot- or cigar-shaped.
The body of the cell is enclosed by a stiff but elastic structure called the pellicle. The pellicle consists of an outer cell membrane (plasma membrane), a layer of flattened membrane-bound sacs called alveoli, and an inner membrane called the epiplasm. The pellicle is not smooth, but textured with hexagonal or rectangular depressions. Each of these polygons is perforated by a central aperture through which a single cilium projects. Between the alveolar sacs of the pellicle, most species of Paramecium have closely spaced spindle-shaped trichocysts, explosive organelles that discharge thin, non-toxic filaments, often used for defensive purposes.[8][9]
Typically, an anal pore (cytoproct) is located on the ventral surface, in the posterior half of the cell. In all species, there is a deep oral groove running from the anterior of the cell to its midpoint. This is lined with inconspicuous cilia which beat continuously, drawing food into the cell.[10] Paramecia are primarily heterotrophic, feeding on bacteria and other small organisms. A few species are mixotrophs, deriving some nutrients from endosymbiotic algae (chlorella) carried in the cytoplasm of the cell.[11]
Osmoregulation is carried out by contractile vacuoles, which actively expel water from the cell to compensate for fluid absorbed by osmosis from its surroundings.[12] The number of contractile vacuoles varies depending on the species.[10]
A Paramecium propels itself by whip-like movements of the cilia, which are arranged in tightly spaced rows around the outside of the body. The beat of each cilium has two phases: a fast "effective stroke," during which the cilium is relatively stiff, followed by a slow "recovery stroke," during which the cilium curls loosely to one side and sweeps forward in a counter-clockwise fashion. The densely arrayed cilia move in a coordinated fashion, with waves of activity moving across the "ciliary carpet," creating an effect sometimes likened to that of the wind blowing across a field of grain.[13]
The Paramecium spirals through the water as it progresses. When it happens to encounter an obstacle, the "effective stroke" of its cilia is reversed and the organism swims backward for a brief time, before resuming its forward progress. This is called the avoidance reaction. If it runs into the solid object again, it repeats this process, until it can get past the object.[14]
It has been calculated that a Paramecium expends more than half of its energy in propelling itself through the water.[15] This ciliary method of locomotion has been found to be less than 1% efficient. This low percentage is nevertheless close to the maximum theoretical efficiency that can be achieved by an organism equipped with cilia as short as those of the members of Paramecium.[16]
Paramecia feed on microorganisms like bacteria, algae, and yeasts. To gather food, the Paramecium makes movements with cilia to sweep prey organisms, along with some water, through the oral groove (vestibulum, or vestibule), and into the cell. The food passes from the cilia-lined oral groove into a narrower structure known as the buccal cavity (gullet). From there, food particles pass through a small opening called the cytostome, or cell mouth, and move into the interior of the cell. As food enters the cell, it is gathered into food vacuoles, which are periodically closed off and released into the cytoplasm, where they begin circulating through the cell body by the streaming movement of the cell contents, a process called cyclosis or cytoplasmic streaming. As a food vacuole moves along, enzymes from the cytoplasm enter it, to digest the contents. As enzymatic digestion proceeds, the vacuole contents become more acidic. Within five minutes of a vacuole's formation, the pH of its contents drops from 7 to 3.[17] As digested nutrients pass into the cytoplasm, the vacuole shrinks. When the fully digested vacuole reaches the anal pore, it ruptures, expelling its waste contents outside the cell.[18][19][20]
Some species of Paramecium form mutualistic relationships with other organisms. Paramecium bursaria and Paramecium chlorelligerum harbour endosymbiotic green algae, from which they derive nutrients and a degree of protection from predators such as Didinium nasutum.[21][22] Numerous bacterial endosymbionts have been identified in species of Paramecium.[23] Some intracellular bacteria, known as kappa particles, give Paramecia the ability to kill other strains of Paramecium that lack kappa particles.[23]
The genome of the species Paramecium tetraurelia has been sequenced, providing evidence for three whole-genome duplications.[24]
In some ciliates, like Stylonychia and Paramecium, only UGA is decoded as a stop codon, while UAG and UAA are reassigned as sense codons (that is, codons that code for standard amino acids), coding for the amino acid glutamic acid.[25]
The question of whether Paramecia exhibit learning has been the object of a great deal of experimentation, yielding equivocal results. However, a study published in 2006 seems to show that Paramecium caudatum may be trained, through the application of a 6.5 volt electric current, to discriminate between brightness levels.[26] This experiment has been cited as a possible instance of cell memory, or epigenetic learning in organisms with no nervous system.[27]
Like all ciliates, Paramecia have a dual nuclear apparatus, consisting of a polyploid macronucleus, and one or more diploid micronuclei. The macronucleus controls non-reproductive cell functions, expressing the genes needed for daily functioning. The micronucleus is the generative, or germline nucleus, containing the genetic material that is passed along from one generation to the next.[28]
Paramecium reproduction is asexual, by binary fission, which has been characterized as "the sole mode of reproduction in ciliates" (conjugation being a sexual phenomenon, not directly resulting in increase of numbers).[2][29] During fission, the macronucleus splits by a type of amitosis, and the micronuclei undergo mitosis. The cell then divides transversally, and each new cell obtains a copy of the micronucleus and the macronucleus.[2]
Fission may occur spontaneously, in the course of the vegetative cell cycle. Under certain conditions, it may be preceded by self-fertilization (autogamy),[30] or it may immediately follow conjugation, in which Paramecia of compatible mating types fuse temporarily and exchange genetic material.
In ciliates such as Paramecium, conjugation is a sexual phenomenon that results in genetic recombination and nuclear reorganization within the cell.[28][23] During conjugation, two Paramecia of a compatible mating type come together and form a bridge between their cytoplasms. Their respective micronuclei undergo meiosis, and haploid micronuclei are exchanged over the bridge. Following conjugation, the cells separate. The old macronuclei are destroyed, and both post-conjugants form new macronuclei, by amplification of DNA in their micronuclei.[28] Conjugation is followed by one or more "exconjugant divisions."[31]
In Paramecium caudatum, the stages of conjugation are as follows (see diagram at right):
In the asexual fission phase of growth, during which cell divisions occur by mitosis rather than meiosis, clonal aging occurs leading to a gradual loss of vitality. In some species, such as the well studied Paramecium tetraurelia, the asexual line of clonally aging Paramecia loses vitality and expires after about 200 fissions if the cells fail to undergo autogamy or conjugation. The basis for clonal aging was clarified by transplantation experiments of Aufderheide in 1986.[32] When macronuclei of clonally young Paramecia were injected into Paramecia of standard clonal age, the lifespan (clonal fissions) of the recipient was prolonged. In contrast, transfer of cytoplasm from clonally young Paramecia did not prolong the lifespan of the recipient. These experiments indicated that the macronucleus, rather than the cytoplasm, is responsible for clonal aging. Other experiments by Smith-Sonneborn,[33] Holmes and Holmes,[34] and Gilley and Blackburn[35] demonstrated that, during clonal aging, DNA damage increases dramatically.[36] Thus, DNA damage in the macronucleus appears to be the cause of aging in P. tetraurelia. In this single-celled protist, aging appears to proceed as it does in multicellular eukaryotes, as described in DNA damage theory of aging.
When clonally aged P. tetraurelia are stimulated to undergo meiosis in association with either conjugation or automixis, the genetic descendants are rejuvenated, and are able to have many more mitotic binary fission divisions. During either of these processes, the micronuclei of the cell(s) undergo meiosis, the old macronucleus disintegrates and a new macronucleus is formed by replication of the micronuclear DNA that had recently undergone meiosis. There is apparently little, if any, DNA damage in the new macronucleus. These findings further solidify that clonal aging is due, in large part, to a progressive accumulation of DNA damage; and that rejuvenation is due to the repair of this damage in the micronucleus during meiosis. Meiosis appears to be an adaptation for DNA repair and rejuvenation in P. tetraurelia.[37] In P. tetraurelia, CtlP protein is a key factor needed for the completion of meiosis during sexual reproduction and recovery of viable sexual progeny.[37] The CtlP and Mre11 nuclease complex are essential for accurate processing and repair of double-strand breaks during homologous recombination.[37]
Paramecium bursaria, a species with symbiotic algae
Paramecium putrinum
Paramecium binary fission
Paramecium in conjugation
Paramecium caudatum
Paramecium aurelia species complex:
Other species:
Paramecium (/ˌpærəˈmiːʃ(i)əm/ PARR-ə-MEE-sh(ee-)əm, /-siəm/ -see-əm; also spelled Paramoecium, plural Paramecia) is a genus of eukaryotic, unicellular ciliates, commonly studied as a model organism of the ciliate group. Paramecia are widespread in freshwater, brackish, and marine environments and are often abundant in stagnant basins and ponds. Because some species are readily cultivated and easily induced to conjugate and divide, they have been widely used in classrooms and laboratories to study biological processes. The usefulness of Paramecium as a model organism has caused one ciliate researcher to characterize it as the "white rat" of the phylum Ciliophora.
Paramecio (O.F. Müller, 1773) estas unuĉela estaĵo, de grupo de grandaj protozooj el la klaso de ciliuloj, ofta en stagna nesala akvo, kiu vivas en riveroj, lagoj, lagetoj kaj akvorestaĵoj. Paramecioj estas travideblaj kaj plataj, kun longeco inter 50 kaj 300 mikrometroj. Ili estas probable la unuĉelaj estaĵoj plej bone konataj kaj la protozooj ciliaj plej studataj de la scienco.
Paramecio (O.F. Müller, 1773) estas unuĉela estaĵo, de grupo de grandaj protozooj el la klaso de ciliuloj, ofta en stagna nesala akvo, kiu vivas en riveroj, lagoj, lagetoj kaj akvorestaĵoj. Paramecioj estas travideblaj kaj plataj, kun longeco inter 50 kaj 300 mikrometroj. Ili estas probable la unuĉelaj estaĵoj plej bone konataj kaj la protozooj ciliaj plej studataj de la scienco.
Los paramecios (género Paramecium) son protistas ciliados con forma ovalada, habituales en aguas dulces estancadas con abundante materia orgánica, como charcos y estanques. Son probablemente los seres unicelulares mejor conocidos y los protozoos ciliados más estudiados por la ciencia. El tamaño ordinario de las especies de paramecios está comprendido entre 0,05 y 0,33 milímetros.
Su superficie está recubierta por cilios muy abundantes, que son heterogéneos tanto en su distribución como en su estructura molecular. A los cilios les corresponde proporcionar información táctil y movimiento al organismo.
El sistema digestivo es simple, con un citostoma externo a largo de la mitad anterior del paramecio, de la que se sirve para capturar el alimento.
Otros orgánulos son el "macronúcleo" eucariota, junto a un "micronúcleo" en el centro del paramecio, y las vacuolas digestivas, que digieren constantemente el alimento capturado. Los desechos se expulsan por exocitosis, mediante vacuolas de secreción que se originan a partir de las digestivas.
Como muchos otros microorganismos, los paramecios se reproducen asexualmente por fisión binaria o mitosis y (sexualmente) por conjugación.

El paramecio es un protozoo de forma ovalada que puede tener una longitud de 50-300 µm en función de la especie.
Los integrantes del género Paramecium muestran un eje anteroposterior permanente y una asimetría izquierda-derecha. Esta polaridad unicelular se refleja en la distribución de estructuras como los cuerpos basales, los cuales se disponen en la cortical en función de aquella.[1]
Toda la superficie de la célula de los Paramecium está recubierta por cilios simples. La mayoría de estos cilios sensoriales, son móviles y están involucrados en dos funciones principales: la alimentación y la locomoción. Los cilios son heterogéneos, tanto en su longitud como en sus propiedades moleculares.[2] Son de pequeña longitud a excepción del surco oral (citostoma) característico, que está recubierto por cilios de mayor longitud y de los cilios de la parte trasera que son más largos que el resto.
Los integrantes de Paramecium son eucariotas que presentan dimorfismo nuclear, caracterizado por la separación de la línea germinal y de la línea somática. Muestran dos tipos de núcleos, un núcleo pequeño llamado "micronúcleo" y otro núcleo grande o "macronúcleo" que suelen ubicarse cerca del centro del organismo.
Según las especies de paramecios se pueden encontrar uno, 2 o 4 micronúcleos. El micronúcleo (MIC) contiene la línea germinal del organismo.[3]
El macronúcleo (MAC) es único y almacena la información somática. Estos núcleos se diferencian desde el núcleo cigótico, después de la fertilización.[4] [5]
En la anatomía externa de los paramecios se destaca el citostoma, una invaginación situada a lo largo del extremo anterior del paramecio, de la que este se sirve para capturar el alimento, conformado por partículas orgánicas flotantes y microorganismos menores. El citostoma conduce a una citofaringe antes de que el alimento pase al interior de este protozoo. Durante las observaciones es relativamente fácil observar la formación de las vacuolas digestivas al final de la citofaringe, su movimiento a través del citoplasma, la disminución de su tamaño al ir digiriéndose su contenido, y la excreción de las partículas no digeridas por el poro anal que se sitúa cerca del extremo trasero.
En algunas especies como Paramecium trichium se observan dos vacuolas contráctiles simples, pero habitualmente se observan vacuolas con canales radiales que les dan un aspecto de estrella muy característico.
Cabe destacar el caso de Paramecium bursaria que vive en simbiosis con algas verdes unicelulares (zooclorelas) que se encuentran libres en su citoplasma y algunas incluso se hallan fijadas en la membrana plasmática, esto les aporta su característico color verde.
La membrana externa absorbe y expulsa regularmente el agua del exterior con el fin de controlar la osmorregulación, proceso dirigido por dos vacuolas contráctiles.
Se alimenta de bacterias, algas y levaduras, ocasionalmente se puede producir la ingesta de protozoos de pequeño tamaño. Para ello se intensifica el movimiento de los cilios situados cerca y dentro del citostoma (boca) creándose de esta manera una corriente de agua y partículas hacia su interior. Las partículas de alimento se van acumulando en la citofaringe y cuando hay suficiente cantidad se genera una vacuola digestiva. Las vacuolas digestivas formadas recorren un circuito específico a través del citoplasma (de la parte posterior a la anterior de la célula) durante el cual son atacadas por enzimas para su digestión, el alimento digerido es absorbido por el citoplasma (se va reduciendo el tamaño de cada una de las vacuolas) y el alimento que llega al final del circuito sin digerir es eliminado a través de un poro anal.
Para facilitar la captura de sus presas cuando el paramecio detecta la presencia de alimento puede disparar tricocistos (orgánulos de forma baciliforme que se sitúan en filas y que se disparan en forma de filamentos terminados en una punta en forma de flecha), este comportamiento también se utiliza como sistema de defensa.
Para compensar el aumento de presión que se produce en la célula como consecuencia de todo este proceso de alimentación, cada célula dispone de dos vacuolas contráctiles que eliminan el exceso de agua continuamente.
Se trata de un protozoo muy común que se encuentra en estanques de agua dulce, acequias, arroyos, ríos, lagos, riachuelos y embalses. En general, su presencia es abundante en aguas estancadas que contienen materia orgánica en descomposición y que, por tanto, son ricas en sustancias nutritivas, como hidratos de carbono y grasa.
Paramecium puede reproducirse de dos maneras distintas. Se puede producir un proceso asexual por división simple (fisión binaria) o se puede producir un proceso de conjugación sexual que tiene lugar en varias fases y que consiste, básicamente, en la unión de dos individuos para el intercambio de su material genético en una secuencia compleja de división, intercambio y fusión de los núcleos. En ambos casos se produce una división transversal del individuo dando lugar a dos células hijas.
En la actualidad se reconocen 12 especies dentro del género Paramecium, aunque existen varias nombradas pero que se consideran dudosas. Las seguras son las siguientes:
Los paramecios (género Paramecium) son protistas ciliados con forma ovalada, habituales en aguas dulces estancadas con abundante materia orgánica, como charcos y estanques. Son probablemente los seres unicelulares mejor conocidos y los protozoos ciliados más estudiados por la ciencia. El tamaño ordinario de las especies de paramecios está comprendido entre 0,05 y 0,33 milímetros.
Su superficie está recubierta por cilios muy abundantes, que son heterogéneos tanto en su distribución como en su estructura molecular. A los cilios les corresponde proporcionar información táctil y movimiento al organismo.
El sistema digestivo es simple, con un citostoma externo a largo de la mitad anterior del paramecio, de la que se sirve para capturar el alimento.
Otros orgánulos son el "macronúcleo" eucariota, junto a un "micronúcleo" en el centro del paramecio, y las vacuolas digestivas, que digieren constantemente el alimento capturado. Los desechos se expulsan por exocitosis, mediante vacuolas de secreción que se originan a partir de las digestivas.
Como muchos otros microorganismos, los paramecios se reproducen asexualmente por fisión binaria o mitosis y (sexualmente) por conjugación.
Kingloom (Paramecium) on ripsloomade klassi ühtlasripsmeliste seltsi kuuluv ainuraksete perekond[1].
Kingloom on väga levinud ainurakne. Kui võtta mõnest tavalisest tiigist natuke vett, siis sisaldab see harilikult kingloomi. Kingloomi on ka lihtne väikestes akvaariumides kasvatada.[2]
Teaduslikult kirjeldas kinglooma esimesena Otto Friedrich Müller aastal 1773.
Kingloom on suhteliselt suur üherakuline organism. Tema keha on 0,1–0,3 mm pikk. Tema välimust ja eluviisi on lihtne jälgida tavalise koolimikroskoobiga.[3]
Nime on ta saanud oma kingakujulise keha järgi.[3] Selline nimi on tal paljudes keeltes: saksa keeles Pantoffeltierchen, hollandi keeles Pantoffeldiertjes, soome keeles Tohvelieläin, vene keeles туфелька.
Kinglooma mass on umbes 1 μg.[4]
Kingloom on pidevas liikumises. Toatemperatuuril on tema liikumiskiirus 2–2½ mm/s, mis on nii pisikese looma kohta suur kiirus: 10–15 kehapikkust sekundis. Edasiliikumise ajal pöörleb ta ümber keha pikitelje paremale.[3]
Tema piklik keha on kaetud lühikeste ripsmetega, mille abil ta liigub edasi. Tal on 10–15 tuhat ripset. Iga ripse teeb kiireid aerulöögitaolisi liigutusi, toatemperatuuril kuni 30 lööki sekundis. Löök on suunatud taha ja selle käigus sirutub ripse täielikult välja. Algseisu tagasi liigub ripse 3–5 korda aeglasemalt ja teeb selle käigus poolkaare. Elektronmikroskoobiga on kindlaks tehtud, et ripsme ehitus on keerukas.[3]
Ripsmete töö on kooskõlastatud. Kõrvuti asuvad ripsmed liiguvad korrapäraste lainetena. Laine algab kinglooma eesosast ja levib tahapoole. Üheaegselt liigub mööda kinglooma keha paar-kolm lainet.
Kinglooma keha eestotsast kuni keskosani kulgeb pikemate ripsmetega sügav lohk – suuväli ehk peristoom. Selle tagumises osas keha keskpaigas on rakusuu, mis viib lühikesse torujasse rakuneelu. Suuvälja ripsmed on jaotunud kahte rühma, mis suunavad lähedusesoleva vee koos selles olevate toiduosakestega neelu. Rakusuu ja neel on pidevalt avatud, kingloom ei saagi suud kinni panna. Kui neelu on kogunenud piisavalt toitaineid, tekib sellest toitevakuool, mis eraldub neelust ja siseneb endoplasmasse.[5]
Kinglooma põhitoiduks on bakterid. Kuid alla neelata võib ta igasuguseid osakesi, sõltumata nende toiteväärtusest. Kingloom on suur õgard: ta sööb vahetpidamata. Üksnes sigimise ajaks teeb ta pausi.[5]
Vakuool ei jää moodustumiskohale. Ta liigub mööda endoplasmat ringi, läbides keerulise ja kindlaksmääratud tee. Seda nimetatakse toitevakuooli tsükloosiks. Tsükloosi käigus seeditakse ära kõik söödav, mis vakuoolis on. See protsess kestab suhteliselt kaua, toatemperatuuril tund aega.[5]
Kingloomal on tüüpiline rakusisene seedimine nagu amööbidel ja mõnel viburloomal. Läbi vakuooli seina tungivad sellesse endoplasmast seedimist soodustavad fermendid. Seeditud toit imendub omakorda läbi vakuooli seina endoplasmasse.[6]
Kui vakuool moodustub, siis on selles vesi, mis ju kinglooma kõikjal ümbritseb. Siis tungivad sellesse aga fermendid ja vakuool muutub tugevalt happeliseks. Selles saab veenduda, lisades toidule happesusindikaatorit, mis vastavalt keskkonna happesusele värvust muudab. Happelises keskkonnas toimub seedimise esimene osa. Seejärel muutub vakuool hoopis nõrgalt leeliseliseks. Sellistes tingimustes toimub ülejäänud seedimine. Tavaliselt kestab seedimise happeline faas kuuendiku kuni veerandi seedimise ajast, aga see võib tunduvalt muutuda, olenedes sellest, mida kingloom sööb.[6]
Seedimata toidujäänused heidetakse välja rakupäraku kaudu. Toitevakuool läheneb kehapinnale, misjärel vakuooli seedimata jäänused ja seda ümbritsev vedelik heidetakse väliskeskkonda. Erinevalt amööbidest, kelle defekatsioon võib toimuda ükskõik millises kehapiirkonnas, juhtub kingloomal see kindlas kehaosas, kõhtmisel küljel suuava ja keha tagaotsa vahelise piirkonna keskel. Kõhtmiseks kehapooleks nimetatakse tinglikult seda kehapoolt, millel asub suuava.[6]
Kingloomal on kaks pulseerivat vakuooli ehk tuikekublikut. Üks neist asub keha eesmises, teine tagumises kolmandikus. Tuikekublik koosneb kesksest reservuaarist ja 5–7 seda radiaalselt ümbritsevast toomakanalist.[7]
Tuikekubliku esimeses tööfaasis koguneb toomakanalitesse vedelik, misjärel kanalid muutuvad hästi nähtavateks, aga keskne reservuaar on siis märkamatu. Teises faasis tühjenevad toomakanalid kesksesse reservuaari, mis muutub hästi märgatavaks, aga toomakanalid seevastu muutuvad märkamatuks. Kolmandas faasis tühjeneb reservuaar läbi erilise rakukestas oleva ava ehk eritusurve ümbritsevasse keskkonda, misjärel pole enam märgata toomakanaleid ega reservuaari. Siis algab protsess uuesti.[7]
Kinglooma esimene ja tagumine tuikekublik töötavad kordamööda. Üks tsükkel kestab 10–15 sekundit. See aga sõltub keskkonnast, sest magevees elavatel kingloomadel töötab tuikekublik hoopis kiiremini kui merevees elavatel ja parasiitsetel kingloomadel. Poole kuni kolmveerand tunniga viiakse tuikekubliku kaudu kehast välja sama suur kogus vedelikku kui on kinglooma ruumala. See vedelik tungib tema kehasse niihästi läbi rakusuu koos toitevakuoolidega kui osmootselt läbi rakuseina.[7]
Kingloom vajab hapnikku 50 ml/kg/h ehk oma kehakaalu kohta 4 korda vähem kui puhkeolekus inimene.[4]
Tuikekubliku abil saab kingloom reguleerida oma keha osmootset rõhku.[7]
Kingloomal on kaks rakutuuma: suurtuum ehk makronukleus ja pisituum ehk mikronukleus. Suurtuum reguleerib kinglooma elutegevust, pisituum osaleb sugulisel paljunemisel. Seda nähtust nimetatakse tuumaaparaadi dualismiks.[7]
Mõlemad tuumad asuvad kinglooma keskel teineteise vahetus naabruses. Väiksem neist on ümaram, suurem on muna- või oakujuline. Struktuurilt on nad erinevad. Suurtuum sisaldab palju rohkem DNA-d ja mitukümmend korda rohkem kromosoome kui pisituum. Suurtuum on omapärane hulgikromosoomne ehk polüploidne tuum.[7]
Kinglooma ektoplasmas on selgesti näha kehapinnaga risti asuvad lühikesed kepikesed, mida nimetatakse paisatiteks ehk trihhotsüütideks. Nad paiknevad korrapäraselt ripsmete vahel, nii et neid on umbes sama palju kui ripsmeid. Need on kinglooma kaitseorganid. Mis tahes ärrituse (sealhulgas keemilise ärrituse) järel viskuvad paisatid välja ja muutuvad pikkadeks niitideks, mis haaravad vaenlast. Kasutatud ehk väljatulistatud paisatite asemele tekivad ektoplasmas uued.[8]
Kingloomadele on omane taksis ehk liikumissuuna ja -kiiruse muutumine väliste ärritajate toimel. Täheldatud on kemotaksist ehk reaktsiooni keemilistele ainetele ja galvanotaksist ehk reaktsiooni elektrivoolule. Taksis võib olla nii positiivne ehk meelitav kui negatiivne ehk heidutav.[3]
Negatiivne taksis on kingloomal keedusoola suhtes. Kui kingloomade elukeskkonda viia keedusoola kristallike, eemalduvad kingloomad sellest kiiresti. Positiivne taksis on neil aga süsihappegaasi suhtes. Kui kingloomade elukohta viia süsihappegaasi mullike, kogunevad kingloomad selle ümber. Kui mulli sees on gaasi kontsentratsioon nii kõrge, et see mõjub kingloomale kahjulikult, jäävad kingloomad sellisele kaugusele, et kontsentratsioon oleks optimaalne, ja moodustavad rõnga.[3]
Kui läbi kingloomi sisaldava vee lasta nõrk elektrivool, siis paigutuvad kõik kingloomad oma pikiteljega paralleelselt elektrivooluga ja hakkavad liikuma katoodi suunas, mille ümber tekib lõpuks tihe kingloomade summ.[3]
Ebasoodsad keskkonnatingimused elab ta üle tsüstina.
Kingloomal on ka vaenlasi. Röövripslooma Didinium nasutum lemmiktoiduks on just kingloomad. Didinium on 0,1–0,15 mm pikk, see tähendab, et kingloomast tüki maad väiksem. Sellegipoolest on Didinium veelgi suurem õgard kui kingloom ise, sest päevas sööb ta keskmiselt 12 kinglooma. Ta on võimeline kinglooma tervenisti alla neelama. Seedimine kestab umbes 2 tundi.[9]
Enamasti paljuneb kingloom mittesugulisel ristpooldumise teel. Pärast aktiivse ujumise ja toitumise perioodi venib kingloom pikemaks ja tema keskkohta tekib ristipidine soon, mis üha enam süveneb. Lõpuks nöördub kingloom pooleks. Tekkivad kaks isendit on geneetiliselt identsed. Kumbki pool saab osa pisituumast ja osa suurtuumast, mis samuti mõlemad poolduvad. Pisituum pooldub varem ja teeb seda mitootiliselt, suurtuum hiljem ja tema pooldumine meenutab väliselt amitoosi. Siis poolduvad ka ripsmejuured ja igast ripsmest tekib kaks, et tütarisenditel oleks ripsmeid sama palju kui emasisendil. Pooldumisprotsess kestab toatemperatuuril tund aega.[10]
Pooldumiselt sarnaneb kingloom üsnagi amööbide ja viburloomadega, vahe on peamiselt selles, et viburloom jaguneb pooleks pikuti ja kingloom põiki.[10]
Aeg-ajalt esineb kingloomadel täiesti eriline sugulise sigimise vorm, mida nimetatakse rakuühteks ehk konjugatsiooniks. Lühidalt toimub see järgmisel viisil. Kaks ripslooma lähenevad teineteisele ja liibuvad tihedalt kõhtudega teineteise vastu. Kummagi kinglooma suurtuum muutub ebastabiilseks ja laguneb, pisituum seevastu pooldub mitu korda, näiteks kaks korda. Saadud 4 pisituumaeksemplarist lahustuvad kõik peale ühe tsütoplasmas. Viimane pisituumaeksemplar pooldub uuesti ja pärast seda liigub saadud pooltest üks teise kinglooma kehasse, teine seevastu jääb paigale. Seejärel on kummaski kingloomas üks pisituumaeksemplar, mis on pärit eri kingloomast. Neid nimetatakse püsituumaks ja siirdtuumaks. Püsituum ja siirdtuum ühinevad omavahel, moodustades ühendtuuma ehk sünkaarüoni. Siiamaale on kaks kinglooma end kogu aeg teineteise vastu surunud ja kõrvuti koos ujunud ning see on aega võtnud kaua aega, isegi 12 tundi. Nüüd võivad nad teineteisest eralduda. Sünkaarüon pooldub viimast korda, ühest poolest saab uus suurtuum ja teisest uus pisituum. Lõpuks lahustuvad vana suurtuuma jäänused tsütoplasmas täielikult.[10]
Rakuühe on vajalik geneetilise mitmekesisuse tekkeks ja suurendamiseks kingloomade asurkonnas, sest pooldumise käigus tekivad üksnes identsed isendid. Päriliku muutlikkuse suurenemine suurendab asurkonna kohastumisvõimet keskkonnaga.[1]
Täheldatud on veel üht nähtust: kauaaegse suguta sigimise ehk pooldumise järel hakkab rakk vananema. Nii pooldumise kui ainevahetuse kiirus väheneb. Seevastu rakuühte järel taastuvad pooldumise ja ainevahetuse algne kiirus. Seetõttu on rakuühtel noorendav mõju ning selle tulemusel tekkinud uusi isendeid võib vaadelda vanemate isendite arvel tekkinud nooremaks põlvkonnaks.[1]
Kõik teadlased ei pea rakuühet sigimise vormiks, sest selle käigus isendite arv ei suurene.[1]
Kingloom (Paramecium) on ripsloomade klassi ühtlasripsmeliste seltsi kuuluv ainuraksete perekond.
Kingloom on väga levinud ainurakne. Kui võtta mõnest tavalisest tiigist natuke vett, siis sisaldab see harilikult kingloomi. Kingloomi on ka lihtne väikestes akvaariumides kasvatada.
Teaduslikult kirjeldas kinglooma esimesena Otto Friedrich Müller aastal 1773.
Paramezioak (Paramecium generoa) ur gezatan bizi diren protozoo ziliodunak dira, itxura obalatua dutenak. 50 eta 300 µm bitarteko luzeera dute, mikroskopio optikoa beharrezkoa izanik beraiek ikusteko.
Ugariak dira ur-putzu, urmael eta ur geldirik duten habitetan.
Aspalditik dira ezagunak protista hauek, lehenengo mikroskopioak asmatu zituztenek (XVII. mendean) "animakulu" txikiak ikusi baitzituzten landareen infusioetan ("infusorio" izena ere eman zieten).
Paramezioek ez dituzte flageloak, zilioak baizik. Zilio horiek protozoo horien azal osoan daude, eta higidura ematen diote organismoari. Paramecium caudatum espezieak 2.000 zilio inguru ditu.
Zapatila itxurako bizidun zelulabakar horiek zitostoma izeneko egitura dute, zelula mintzaren inbaginazioa, aho moduan jarduten duena elikagaiak harrapatu eta irensteko. Zitostomaz gain bi nukleo dituzte, bata bestearen ondoan: nukleo handia (makronukleoa) eta nukleo txikia (mikronukleoa).
Bakterioez eta partikula organiko txikiez elikatu ohi dira. Zilioek ur korrenteak sortzen dituzte bakterioak zitostomara erakartzeko. Irentsitako bakterioak elikagai-bakuola batean pilatzen dira.
Ugalketari dagokionez erdibiditzearen bidez sortzen dira bi paramezio, paramezio-ama batengandik. Ugalketa asexual horretan sortutako paramezioek ama-zelularen ondare genetiko bera dute. Ugalketa sexuala ere egin dezakete: bi paramezio elkartu eta mikronukleoak elkartrukatzen dituzte, material genetiko berria agertuz trukaketaren ondorioz. Ugalketa sexual honi konjugazioa deritzo, eta ugalketa asexualaren aldean urriagoa da.
Paramecium generoan espezie asko daude. Besteak beste, Paramecium caudatum, Paramecium aurelia eta Paramecium bursaria.
Paramezioak (Paramecium generoa) ur gezatan bizi diren protozoo ziliodunak dira, itxura obalatua dutenak. 50 eta 300 µm bitarteko luzeera dute, mikroskopio optikoa beharrezkoa izanik beraiek ikusteko.
Ugariak dira ur-putzu, urmael eta ur geldirik duten habitetan.
Aspalditik dira ezagunak protista hauek, lehenengo mikroskopioak asmatu zituztenek (XVII. mendean) "animakulu" txikiak ikusi baitzituzten landareen infusioetan ("infusorio" izena ere eman zieten).
Paramezioek ez dituzte flageloak, zilioak baizik. Zilio horiek protozoo horien azal osoan daude, eta higidura ematen diote organismoari. Paramecium caudatum espezieak 2.000 zilio inguru ditu.
Zapatila itxurako bizidun zelulabakar horiek zitostoma izeneko egitura dute, zelula mintzaren inbaginazioa, aho moduan jarduten duena elikagaiak harrapatu eta irensteko. Zitostomaz gain bi nukleo dituzte, bata bestearen ondoan: nukleo handia (makronukleoa) eta nukleo txikia (mikronukleoa).
Bakterioez eta partikula organiko txikiez elikatu ohi dira. Zilioek ur korrenteak sortzen dituzte bakterioak zitostomara erakartzeko. Irentsitako bakterioak elikagai-bakuola batean pilatzen dira.
Ugalketari dagokionez erdibiditzearen bidez sortzen dira bi paramezio, paramezio-ama batengandik. Ugalketa asexual horretan sortutako paramezioek ama-zelularen ondare genetiko bera dute. Ugalketa sexuala ere egin dezakete: bi paramezio elkartu eta mikronukleoak elkartrukatzen dituzte, material genetiko berria agertuz trukaketaren ondorioz. Ugalketa sexual honi konjugazioa deritzo, eta ugalketa asexualaren aldean urriagoa da.
Paramecium generoan espezie asko daude. Besteak beste, Paramecium caudatum, Paramecium aurelia eta Paramecium bursaria.
Tohvelieläimet (Paramecium) ovat ripsieläimiin kuuluvia yksisoluisia eliöitä. Ne kuuluvat Chromalveolata-eliökuntaan. Muiden ripsieläinten tavoin tohvelieläimillä on solukalvossa ripsiä, joiden avulla solu liikkuu ja pyydystää ravintohiukkasia. Tohvelieläimet ovat 100–300 mikrometriä pitkiä, joten niitä ei näe paljain silmin, mutta sen voi nähdä liikehtivän vesilätäkössä. Ravinnokseen ne käyttävät sekä erilaisia bakteereja että muita pienempiä yksisoluisia eliöitä ja kasvisoluja.
Tohvelieläimet (Paramecium) ovat ripsieläimiin kuuluvia yksisoluisia eliöitä. Ne kuuluvat Chromalveolata-eliökuntaan. Muiden ripsieläinten tavoin tohvelieläimillä on solukalvossa ripsiä, joiden avulla solu liikkuu ja pyydystää ravintohiukkasia. Tohvelieläimet ovat 100–300 mikrometriä pitkiä, joten niitä ei näe paljain silmin, mutta sen voi nähdä liikehtivän vesilätäkössä. Ravinnokseen ne käyttävät sekä erilaisia bakteereja että muita pienempiä yksisoluisia eliöitä ja kasvisoluja.
Paramecium
La Paramécie, Paramecium, est un genre d'organismes eucaryotes unicellulaires (cellule ayant un noyau et des organites), ou protistes, dont certaines espèces (en particulier P. caudatum) sont couramment utilisées comme organisme modèle dans les laboratoires de microbiologie. Elles font partie de l'embranchement des ciliés, dans la division des alvéolés[1].
Le terme paramécie est issu de l'adjectif grec ancien παραμήκης qui signifie oblong, allongé, qui s'étend en longueur.
On les élevait autrefois sous forme d'infusoires pour les besoins scolaires (découverte des micro-organismes en sciences naturelles) ou pour nourrir les alevins de jeunes poissons (aquariophilie).
La taille de la cellule varie de 0,1 à 0,3 mm de long suivant les espèces.
La paramécie utilise des cils pour se déplacer et se nourrir (essentiellement de bactéries, par phagocytose).
Les paramécies sont aussi caractérisées par une des exceptions à l'universalité du code génétique (avec les mitochondries).
Les paramécies vivent isolées en eau douce, mais peuvent produire des biofilms caractéristiques quand leur nourriture est abondante (généralement en contexte d'eutrophisation locale et momentanée due à la décomposition de feuilles mortes). Elles peuvent être trouvées en abondance dans les étangs, de petits cours d'eau lents, et en biofilm en surface de flaques ou d'eaux stagnantes (en présence de végétaux morts).
Elles font partie des « infusoires » des anciens auteurs, nom qui provient du fait qu'elles apparaissent en grand nombre dans les infusions de végétaux (foins, thé, peaux de fruits, etc.), rendant leur culture et leur étude aisées.
Vivant dans un milieu hypotonique par rapport à son cytoplasme, la cellule absorbe constamment l'eau de son environnement par osmose. L'excès d'eau dans le cytoplasme est alors évacué grâce à des vacuoles pulsatiles, où le cytoplasme se contracte périodiquement pour expulser l'eau à travers la membrane plasmique. (Un pore est mis en place au-dessus de la vacuole en systole, ce de manière temporaire.)
Comme la plupart des ciliés, la paramécie présente la particularité d'avoir un appareil nucléaire en deux parties : un (ou plusieurs) petit noyau, le micronucleus, et un gros noyau, le macronucleus. Le premier assure les fonctions sexuelles indispensables pour engendrer des variations génétiques alors que le second dirige les fonctions trophiques quotidiennes et la multiplication asexuée, c'est un organisme qui est diplobionte (cycle de reproduction diplobiontique). La reproduction de la paramécie est asexuée, elle se fait par scissiparité.
Pour survivre, la paramécie effectue des échanges de matière par diffusion avec le milieu extérieur. La digestion se fait dans la cellule, grâce à un système de vacuoles spécialisées.
L'histoire scientifique des paramécies se confond avec celle des infusoires, qui ont fait partie des espèces microscopiques unicellulaires les plus étudiées depuis le XVIIIe siècle, après l'invention du microscope et quand les hygiénistes ont rendu les microbes particulièrement intéressants pour le grand public, les enseignants, les élèves et les scientifiques.
Elles sont l'un des premiers organismes à avoir été observé au microscope.
La première description de paramécie date du XVIIIe siècle (Fig. 1).
Les premiers observateurs croient à une génération spontanée, ou pensent que les végétaux morts se décomposent en ces petits animalcules. Certains croient voir des accouplements ou des œufs, des vésicules séminales ou des ovaires (qui sont en fait des vacuoles que l'on peut colorer) et des pontes (probablement des « exudations de sarcodes »[2] ou vacuoles perdues par des organismes morts). Ces erreurs seront peu à peu corrigées et l'étude des infusoires contribuera à fournir des preuves de plus de la validité de l'axiome Omne vivum ex ovo (inexistence de la génération spontanée). Différents auteurs ont observé des infusions de foin, de blé, d'avoine, de poivre, de thé, etc. et cherché à distinguer les infusoires d'autres catégories d'animalcules (brachions notamment).
En 1744, Trembley découvre que certains infusoires peuvent parasiter ou coloniser d'autres organismes tels que l'hydre d'eau douce et décrit aussi des organismes « vorticelliens » en distinguant dans ce groupe ce qu'il nomme « polypes à bulbes » et « polype à bras »[3].
En 1752, dans un essai sur l'histoire naturelle, Hill commence à chercher à donner des noms scientifiques aux infusoires, mais sa tentative de classification sera rapidement oubliée[4]
En 1754, Joblot se passionne pour les microorganismes aquatiques et produit quelques bonnes descriptions à partir du microscope, mais assorties à des dénominations personnelles et des commentaires fantaisistes, et en mélangeant les infusoires avec d'autres groupes d'espèces[5].
En 1764, Wrisberg publie des observations sur la nature des animalcules infusoires (1re dénomination de ce type)
En 1758 (puis 1767), Linné qui ne les a pas étudiés par lui-même les confond d'abord dans Systema naturae (Ed. X, 1758 et Ed XII, 1767) sous le terme trop imprécis de Chaos, en distinguant cependant le Volvox globator, puis en admettant un genre Vorticella.
En 1766, Pallas publie un ouvrage sur les zoophytes où il réunit seulement dans deux genres( Volvox et Brachionus) les animalcules microscopiques qu'il connait[6].
En 1769, Ellis décrit plusieurs infusoires sous le nom de Volvox (transactions philosophiques de Londres, 1769[7], mais c'est surtout le naturaliste danois Otto-Frédéric Müller qui après avoir travaillé sur une classification des vers marins et fluviatiles[8] tente de méthodiquement classer les infusoires selon la méthode linnéenne (grâce à un microscope composé). Puis après l'apparition du microscope achromatique et sur la base des découvertes et hypothèse de Christian Gottfried Ehrenberg dès 1830, la classification et l'organisation des infusoires est précisée. Cette même année, Eichhorn - sans les classer (il se borne à leur donner des noms descriptifs en allemand) - en décrit un nombre plus important que tous ses prédécesseurs. Il présente aussi dans ce groupe des organismes qui ne sont pas des infusoires[9]. Les derniers travaux de Müller sont publiés en 1786[10] à titre posthume (notes mises en forme par Fabricius), mais incomplets et non révisés par l'auteur, mort avant d'avoir terminé ce travail. À cette époque, la classification des 379 espèces d'infusoires décrits par Müller, classés par lui en 17 genres est encore très imparfaite avec de nombreux doublons, des descriptions textuelles peu claires (mais appuyées sur des représentations graphiques de plus en plus précises), et des incertitudes taxonomiques pour environ la moitié des espèces alors décrites. Félix Dujardin admire son travail, mais lui reproche, comme à ses prédécesseurs, d'avoir encore confondu des « propagules d'algues, des Bacillaires, des Navicules, des Anguillules, des Distomes, de jeunes Alcyonelles, des lambeaux de branchies de mollusques » avec des infusoires et d'avoir plusieurs fois confondu des restes incomplets d'organismes, ou des états différents d'un même organisme avec une espèce en tant que telle[9].
En 1776, Spallanzani étudie plus spécifiquement quelques infusoires et les rotifères, et avec son ami Saussure publie quelques-unes de ses découvertes sur ces organismes[9].
En 1778, Gleichen s'y intéresse dans ses travaux sur la génération des êtres vivants, mais les décrit piètrement[11].
En 1782, Goeze[12] et Bloch[13], séparément, en étudiant les vers intestinaux découvrent des infusoires vivant dans l'intestin des grenouilles ;
Cuvier, comme Linné s'est peu intéressé aux infusoires et à leur classification (il en a séparé en 1817, à tort selon ....... « les vraies Vorticelle qu'il plaçait dans son ordre des polypes gélatineux ; et il avait senti la nécessité de séparer les Systolides pourvus d'un intestin et d'organes compliqués, et les vrais Infusoires, "animaux à corps gélatineux de la plus extrême simplicité, sans viscères, et souvent même sans une apparence de bouche" »[14].
Lamarck, dans son Histoire des animaux sans vertèbres (1815-1819)[15] conserve à tort une partie erronée de la classification de Müller ; tout en démembrant à raison plusieurs de ses genres dont celui des Vorticelles d'où il retira les Rotifères et les autres Systolides pour en faire son genre Furculaire ; mais « n'ayant point observé par lui-même, il laissa subsister dans les divers genres les autres rapprochements erronés de Müller, et même en ajouta de nouveaux dans son genre Furcocerque. Il plaça avec raison les Systolides dans une autre classe que les Infusoire‘s proprement dits, mais avec eux, il eut le tort de placer les Vorticelles parmi les Polypes ciliés » note Dujardin (1841)[9].
En 1817, l'allemand Nitzsch améliore la connaissance des Navicules et des Cercaires qu’il démontre n’être pas de vrais Infusoires.
En 1825, Bory de Saint-Vincent, alors chargé de terminer la partie de l'Encyclopédie méthodique commencée par Bruguières, est invité à vérifier la classification des Infusoires (qu'il veut renommer des « Microscopiques » (...)« d’après cette considération que beaucoup d'entre eux vivent dans les eaux pures et non dans les infusions »[9]).
Il subdivise à cette occasion les 17 genres établis par Müller en 99 genres, plus précis dont certains valent encore, mais dans sa classe des Microscopiques, « il laisse encore confondus les Systolides, et il en distrait les seules Vorticelles pédicellées qu’il reporte, avec les Navicules et les Lunulines, dans son règne psychodiaire »[9].
En 1827, dans une Encyclopédie allemande Nitzsch propose la création de nouveaux genres (« M. Dutrochet, en France, avait étudié les Rotifères et les Tubicolaires; M. Leclerc avait fait connaître les Difflugies; et Losana , en Italie , avait décrit des Amibes, des Kolpodes et des Cyclides dont il multipliait les espèces sans raison et sans mesure », selon Dujardin (1841)[9]).
En 1830, Christian Gottfried Ehrenberg renomme ensuite le groupe des Infusoires en Polygastrica (car il estime - à tort - qu'ils ont une bouche et plusieurs estomacs), il en extrait les Systolides (qu’il nomme Rotatoria) mais conserve comme vrais Infusoires à la fois les Clostéries ou Lunulines, les Navicules et toutes les Diatomés et Desmidiées, ce qui lui permet de porter le nombre des Infusoires (ou polygastriques) à 533[9]. Quelques auteurs et compilateurs reprennent cette nouvelle classification, mais elle est rapidement contestée[9].
Des botanistes allemands et français forcent à ensuite classer les Navicules et les Clostéries dans le règne végétal[9].
En 1841, le naturaliste Félix Dujardin publie une histoire naturelle des zoophytes incluant une description de la physiologie et une classification des infusoires, ainsi qu'une présentation de « la manière de les étudier à l'aide du microscope » [9].
La classification en espèces est toujours en cours — par exemple, P. aurelia a récemment été divisée en 14 espèces — mais la classification suivante est représentative :
Paramecium
La Paramécie, Paramecium, est un genre d'organismes eucaryotes unicellulaires (cellule ayant un noyau et des organites), ou protistes, dont certaines espèces (en particulier P. caudatum) sont couramment utilisées comme organisme modèle dans les laboratoires de microbiologie. Elles font partie de l'embranchement des ciliés, dans la division des alvéolés.
Le terme paramécie est issu de l'adjectif grec ancien παραμήκης qui signifie oblong, allongé, qui s'étend en longueur.
On les élevait autrefois sous forme d'infusoires pour les besoins scolaires (découverte des micro-organismes en sciences naturelles) ou pour nourrir les alevins de jeunes poissons (aquariophilie).
Prótasón, speiceas mionorgánaigh aoncheallaigh, ubhchruthach, suas le .33 mm ar fhad. Dhá núicléas atá ann, micreanúicléas is macranúicléas. Itheann sé baictéir. Go cothrom ar fud an dromchla tá fabhráin, cosúil le ribí gruaige. An-choiteann i ngnáthóga uisciúla.
Os paramecios son protozoos ciliados do xénero Paramecium, que viven en augas doces estancadas con abundancia de materia orgánica. Son uns protozoos ovalados [1] de 50 a 350 microns de lonxitude segundo a especie, probablemente uns dos seres unicelulares mellor coñecidos e máis estudados. No se deben confundir con Blepharisma, que é un xénero parecido.
Teñen numerosos cilios que cobren toda a súa superficie e que lle dan mobilidade, os da parte posterior algo maiores. Tamén ten cilios no citostoma, que son un pouco máis grandes e lle axudan a capturar o alimento. Para moverse cara a adiante os seus cilios baten nun ángulo de 120 graos, cara atrás todos ao unísono. O paramecio móvese facendo espirais pola auga. Pode moverse tamén para atrás se os cilios baten formando un ángulo cara a adiante todos á vez. Se choca contra un obxecto recúa, xírase lixeiramente e vai de novo para adiante, repetindo isto ata que salva o obstáculo. Pode moverse nun segundo unha distancia 12 veces maior que o seu tamaño.
A membrana externa absorbe e expulsa regularmente auga do exterior coa fin de controlar a osmorregulación, proceso dirixido por dous vacúolos contráctiles.[2] Está recuberto por unha película, que é unha membrana resistente pero elástica.
Nalgunhas especies como P. trichium hai dous vacúolos contráctiles simples, pero o máis habitual é que estes vacúolos teñan unhas estriacións radiais que lle dan un aspecto de estrela.
Presentan un citostoma, unha especie de invaxinación situada a todo o longo do paramecio coa que captura o alimento, que consiste en partículas orgánicas flotantes e microorganismos menores ca el. O citostoma conduce a unha citofarinxe antes de que o alimento pase ao interior deste protozoo. Son moi visibles os vacúolos dixestivos, onde dixiren o alimento, e despois expulsan por exocitose os refugallos.
Presentan polo menos dous núcleos de distinto tamaño, chamados micronúcleo e macronúcleo. O macronúcleo está no centro e é de gran tamaño e forma alongada, compacto, poliploide, con funcións vexetativas, e o único que transcribe. O micronúcleo é pequeno e máis esférico (en P. aurelia hai dous micronúcleos), non transcribe ARN, pero é o que realiza a meiose e a conxugación.
Poden reproducirse por fisión binaria (mitose) ou por conxugación entre dous individuos. En P. caudatum durante a conxugación únense dous individuos, dexeneran os macronúcleos, os micronúcleos sofren meiose, formando 4 núcleos cada un deles (hai 8 entre os dous conxugates) dos cales dexeneran 3, e os outros dous que quedan divídense unha vez (agora son catro) e fusiónanse cos do outro individuo, formando dous núcleos, un en cada individuo, momento no que os individuos se separan. Unha vez separados hai máis divisións nucleares nas que se rexenera o macronúcleo e o micronúcleo iniciais. Como resultado da meiose e da conxugación houbo unha recombinación e intercambio de xenes.[3] Na conxugación non se unen organismos completos (o que sería singamia) senón só os seus núcleos.
Os paramecios aliméntanse de bacterias, algas e lévedos, e ocasionalmente pódese producir a inxesta de protozoos de pequeno tamaño. Pode detectar quimiotacticamente metabolitos bacterianos presentes no seu medio, e ten receptores superficiais para o folato, acetato e glutamato. Para capturalos incrementa o movemento dos cilios situados arredor e dentro do citostoma creando unha corrente de auga e partículas cara ao seu interior. As partículas de alimento vanse acumulando na citofarinxe e cando hai suficiente cantidade fórmase un vacúolo dixestivo. Os vacúolos dixestivos formados percorren un circuíto específico a través do citoplasma (da parte posterior á anterior da célula) durante o cal o seu contido se dixire encimaticamente, e o alimento dixerido absórbese cara ao citoplasma mentres o vacúolo reduce o seu tamaño. Os restos sen dixerir, cando se chega ao final do circuíto, son expulsados a través dun poro anal.
Para facilitar a captura das súas presas Paramecium pode disparar tricocistes, orgánulos de forma baciliforme que se sitúan en ringleiras e que se disparan en forma de filamentos rematados en punta de frecha e segregan un produto ; este comportamento tamén se utiliza como sistema de defensa.
É un protozoo moi común que se encontra en pozas e estanques de auga doce, cales, regatos, ríos, lagos e encoros. En xeral a súa presenza é abundante en augas estancadas que conteñen materia orgánica en descomposición e que, por tanto, son ricas en substancias nutritivas e abundantes en microorganismos. Recentemente, informouse de novas especies de paramecios atopadas en augas salgadas.[4]
O xenoma de Paramecium foi secuenciado (na especie Paramecium tetraurelia), e presenta evidencias de tres duplicacións de todo o xenoma.[5]
Tamén presenta algunhas variacións con respecto ao código xenético estándar. En Paramecium (e noutros ciliados como Stylonychia), só hai un codón de parada da tradución de proteínas, que é UGA (normalmente hai tres), entanto que os codóns UAG e UAA, que noutros seres son tamén de parada, nos paramecios están reasignados como codóns con sentido que codifican aminoácidos.[6]
No Paramecium bursaria prodúcese un caso de endosimbiose. Este protozoo vive en simbiose con algas verdes unicelulares do xénero Chlorella, que se encontran no seu citoplasma e lle dan cor verde ao protozoo [7]. Ademais, P. aurelia ten endosimbiontes bacterianos.[8]
Na actualidade recoñécense 14 especies dentro do xénero Paramecium, aínda que foron nomeadas máis, que se consideran dubidosas. Os estudos xenéticos están restruturando a clasificación deste xénero. As principais especies son:
Os paramecios son protozoos ciliados do xénero Paramecium, que viven en augas doces estancadas con abundancia de materia orgánica. Son uns protozoos ovalados de 50 a 350 microns de lonxitude segundo a especie, probablemente uns dos seres unicelulares mellor coñecidos e máis estudados. No se deben confundir con Blepharisma, que é un xénero parecido.
Teñen numerosos cilios que cobren toda a súa superficie e que lle dan mobilidade, os da parte posterior algo maiores. Tamén ten cilios no citostoma, que son un pouco máis grandes e lle axudan a capturar o alimento. Para moverse cara a adiante os seus cilios baten nun ángulo de 120 graos, cara atrás todos ao unísono. O paramecio móvese facendo espirais pola auga. Pode moverse tamén para atrás se os cilios baten formando un ángulo cara a adiante todos á vez. Se choca contra un obxecto recúa, xírase lixeiramente e vai de novo para adiante, repetindo isto ata que salva o obstáculo. Pode moverse nun segundo unha distancia 12 veces maior que o seu tamaño.
A membrana externa absorbe e expulsa regularmente auga do exterior coa fin de controlar a osmorregulación, proceso dirixido por dous vacúolos contráctiles. Está recuberto por unha película, que é unha membrana resistente pero elástica.
Nalgunhas especies como P. trichium hai dous vacúolos contráctiles simples, pero o máis habitual é que estes vacúolos teñan unhas estriacións radiais que lle dan un aspecto de estrela.
Presentan un citostoma, unha especie de invaxinación situada a todo o longo do paramecio coa que captura o alimento, que consiste en partículas orgánicas flotantes e microorganismos menores ca el. O citostoma conduce a unha citofarinxe antes de que o alimento pase ao interior deste protozoo. Son moi visibles os vacúolos dixestivos, onde dixiren o alimento, e despois expulsan por exocitose os refugallos.
Presentan polo menos dous núcleos de distinto tamaño, chamados micronúcleo e macronúcleo. O macronúcleo está no centro e é de gran tamaño e forma alongada, compacto, poliploide, con funcións vexetativas, e o único que transcribe. O micronúcleo é pequeno e máis esférico (en P. aurelia hai dous micronúcleos), non transcribe ARN, pero é o que realiza a meiose e a conxugación.
Papučica je heterotrofni jednostanični organizam.
Otkrio ju je 1675. godine nizozemski prirodoslovac Antoni van Leeuwenhoek pod svojim jednostavnim svjetlosnim mikroskopom. Kao i svi trepetljikaši, vrlo je rasprostranjena u morima, kopnenim vodama, bočatim vodama i na vlažnom tlu.
Zbog svog oblika je dobila današnji naziv. Razlikuje se od euglene po tome što ima veliku i malu staničnu jezgru. Tijelo joj je obavijeno prozirnom membranom zvanom pelikula, na kojoj se nalaze brojne trepetljike koje joj služe za kretanje. Slično kao i ameba, diše difuzijom plinova preko površine tijela.
Na pelikuli se nalazi jedno mjesto na kojem su trepetljike duže, pa formiraju udubljenje koje nazivamo usno predvorje, te završava staničnim ustima ("citostom") pomoću kojega papučica prima hranjive tvari. Stanično ždrijelo se nastavlja na stanična usta i na njegovom se kraju stvaraju hranidbeni mjehurići. Oni se spajaju s lizosomima koji razgrađuju hranu, a neprobavljeni ostaci se izbacuju na određenom mjestu na membrani, tzv. stanični izmetni otvor.
Papučica se razmnožava diobom stanica. Kad naiđu nepovoljni uvjeti,papučica se kristalizira. Tada se ona ne hrani i ne razmnožava. Kada prođu nepovoljni uvjeti, ona nastavlja normalan život. Nakon više uzastopnih dioba, kad stanica postigne najmanju veličinu, dolazi do stapanja dviju papučica. Nastupa obnavljanje jezgri, te se nakon toga papučice razdvoje. Iz ovog procesa se ne razvija potomstvo, ali ga zovemo spolno razmnožavanje.
Papučica je heterotrofni jednostanični organizam.
Otkrio ju je 1675. godine nizozemski prirodoslovac Antoni van Leeuwenhoek pod svojim jednostavnim svjetlosnim mikroskopom. Kao i svi trepetljikaši, vrlo je rasprostranjena u morima, kopnenim vodama, bočatim vodama i na vlažnom tlu.
Paramecium merupakan salah satu protista mirip hewan. Protista ini berukuran sekitar 50-350ɰm. Paramecium telah memiliki selubung inti (Eukariot). Uniknya Protista ini memiliki dua inti dalam satu sel, yaitu inti kecil (Mikronukleus) yang berfungsi untuk mengendalikan kegiatan reproduksi, dan inti besar (Makronukleus) yang berfungsi untuk mengawasi kegiatan metabolisme, pertumbuhan, dan regenerasi. Paramecium bereproduksi secara aseksual (membelah diri dengan cara transversal), dan seksual (dengan konjugasi). Paramecium bergerak dengan menggetarkan silianya. Hal ini akan terlihat jika menggunakan mikroskop. Mereka menangkap makanan dengan cara menggetarkan silianya, maka terjadi aliran air keluar dan masuk mulut sel. Saat itulah bersamaan dengan air masuk bakteri bahan organik atau hewan uniseluler lainnya. memiliki vakuola makanan yang berfungsi untuk mencerna dan mengedarkan makanan, serta vakuola berdenyut yang berguna untuk mengeluarkan sisa makanan. Kegunaannya sebagai organisme model telah menyebabkan salah satu peneliti Ciliata untuk menggambarkannya sebagai "tikus putih" dari filum Ciliophora.[1]
Paramecium merupakan salah satu protista mirip hewan. Protista ini berukuran sekitar 50-350ɰm. Paramecium telah memiliki selubung inti (Eukariot). Uniknya Protista ini memiliki dua inti dalam satu sel, yaitu inti kecil (Mikronukleus) yang berfungsi untuk mengendalikan kegiatan reproduksi, dan inti besar (Makronukleus) yang berfungsi untuk mengawasi kegiatan metabolisme, pertumbuhan, dan regenerasi. Paramecium bereproduksi secara aseksual (membelah diri dengan cara transversal), dan seksual (dengan konjugasi). Paramecium bergerak dengan menggetarkan silianya. Hal ini akan terlihat jika menggunakan mikroskop. Mereka menangkap makanan dengan cara menggetarkan silianya, maka terjadi aliran air keluar dan masuk mulut sel. Saat itulah bersamaan dengan air masuk bakteri bahan organik atau hewan uniseluler lainnya. memiliki vakuola makanan yang berfungsi untuk mencerna dan mengedarkan makanan, serta vakuola berdenyut yang berguna untuk mengeluarkan sisa makanan. Kegunaannya sebagai organisme model telah menyebabkan salah satu peneliti Ciliata untuk menggambarkannya sebagai "tikus putih" dari filum Ciliophora.
Il Paramecium (O.F.Müll., 1773) è un genere di protisti infusori (così chiamati perché viventi in acque un tempo ricche di fiori e fieno in infusione). Nella classificazione tradizionale vengono considerati protozoi appartenenti alla classe dei ciliati, ordine degli oligotrichi, di cui sono i rappresentanti più tipici. Nelle classificazioni più moderne i parameci sono considerati protisti appartenenti ai ciliofori e nell'ordine dei peniculidi. Si caratterizza per la sua forma ovale, la cui superficie è ricoperta da ciglia che permettono lo spostamento. Date le sue dimensioni, per poterlo osservare è necessario l'utilizzo di un microscopio.
I parameci vivono in acqua dolce[1], stagnante, quindi in un ambiente con concentrazione di soluti molto più bassa di quella interna alla cellula. Per contrastare l'ingresso di acqua per osmosi, il paramecio ha sviluppato una rete di collettori, che portano acqua a una riserva centrale, chiamata vacuolo contrattile, da cui l'acqua è pompata nuovamente nell'ambiente circostante.
Si muove velocemente nell'acqua grazie al battito di numerosissime ciglia di cui sono rivestiti in maniera pressoché uniforme[1], seguendo una traiettoria dall'andamento a spirale, ruotando rispetto all'asse maggiore. Se incontra un ostacolo, mostra la cosiddetta "reazione di esitamento", indietreggiando in diagonale e ripartendo in una nuova direzione.
I batteri costituiscono la sua principale fonte di nutrimento[1]: essi vengono introdotti all'interno del paramecio attraverso un'apertura della membrana cellulare, il citostoma, circondato da ciglia; poi attraverso un'altra apertura, il citopigio, vengono eliminate le sostanze di rifiuto. I parameci, a loro volta, costituiscono la principale fonte di nutrimento per la maggior parte delle specie del genere di ciliati Didinium.
Nella cellula si trovano vacuoli digestivi e vacuoli pulsanti per l'eliminazione dell'acqua in eccesso.
Il paramecio è dotato di micronuclei che controllano la riproduzione e di macronuclei che controllano le altre funzioni. Si riproduce asessualmente per mitosi, ma ottiene la ricombinazione grazie alla coniugazione.
I parameci possiedono anche la possibilità di ospitare al loro interno dei cloroplasti. Infatti, mangiando essi alghe e vegetali in genere, i cloroplasti non vengono distrutti, bensì conservati all'interno del protista; qui svolgono la loro abituale attività di fotosintesi clorofilliana e le sostanze derivanti da questo processo vanno a beneficio del paramecio. Quindi si può dire che, quando un paramecio ospita dei cloroplasti al suo interno, diventa un organismo autotrofo, o meglio un organismo autotrofo facoltativo.
Il Paramecium (O.F.Müll., 1773) è un genere di protisti infusori (così chiamati perché viventi in acque un tempo ricche di fiori e fieno in infusione). Nella classificazione tradizionale vengono considerati protozoi appartenenti alla classe dei ciliati, ordine degli oligotrichi, di cui sono i rappresentanti più tipici. Nelle classificazioni più moderne i parameci sono considerati protisti appartenenti ai ciliofori e nell'ordine dei peniculidi. Si caratterizza per la sua forma ovale, la cui superficie è ricoperta da ciglia che permettono lo spostamento. Date le sue dimensioni, per poterlo osservare è necessario l'utilizzo di un microscopio.
I parameci vivono in acqua dolce, stagnante, quindi in un ambiente con concentrazione di soluti molto più bassa di quella interna alla cellula. Per contrastare l'ingresso di acqua per osmosi, il paramecio ha sviluppato una rete di collettori, che portano acqua a una riserva centrale, chiamata vacuolo contrattile, da cui l'acqua è pompata nuovamente nell'ambiente circostante.
Si muove velocemente nell'acqua grazie al battito di numerosissime ciglia di cui sono rivestiti in maniera pressoché uniforme, seguendo una traiettoria dall'andamento a spirale, ruotando rispetto all'asse maggiore. Se incontra un ostacolo, mostra la cosiddetta "reazione di esitamento", indietreggiando in diagonale e ripartendo in una nuova direzione.
I batteri costituiscono la sua principale fonte di nutrimento: essi vengono introdotti all'interno del paramecio attraverso un'apertura della membrana cellulare, il citostoma, circondato da ciglia; poi attraverso un'altra apertura, il citopigio, vengono eliminate le sostanze di rifiuto. I parameci, a loro volta, costituiscono la principale fonte di nutrimento per la maggior parte delle specie del genere di ciliati Didinium.
Nella cellula si trovano vacuoli digestivi e vacuoli pulsanti per l'eliminazione dell'acqua in eccesso.
Il paramecio è dotato di micronuclei che controllano la riproduzione e di macronuclei che controllano le altre funzioni. Si riproduce asessualmente per mitosi, ma ottiene la ricombinazione grazie alla coniugazione.
I parameci possiedono anche la possibilità di ospitare al loro interno dei cloroplasti. Infatti, mangiando essi alghe e vegetali in genere, i cloroplasti non vengono distrutti, bensì conservati all'interno del protista; qui svolgono la loro abituale attività di fotosintesi clorofilliana e le sostanze derivanti da questo processo vanno a beneficio del paramecio. Quindi si può dire che, quando un paramecio ospita dei cloroplasti al suo interno, diventa un organismo autotrofo, o meglio un organismo autotrofo facoltativo.
Paramecium (O.F. Müller, 1773) est animans unicellularis, soleaeformis, habitat flumina, lacus, stagna et lacunae tenues (turbidesque), ideo pars oecosystematis magno momenti. Paramecia sunt de 0,05 ad 0,32mm longa (species permagnae videri possumus etiam nudis oculis). Praedam (ut bacteria) per chemoreceptores indagatam ciliorum motu sumitur et per cystostoma in vacuolam cibariam defertur (quod endocytosim dicamus). Concoctionis reliquiae per cyclosim trans cellulam deferuntur et extra feruntur per cytopygem (quod exocytosim dicitur). Amplificatio asexualiter per divisionem ex transverso aut et sexualiter per coniugationem (cystostomatibus coniungetibus qua data hereditiaria traduntur). Motus in aqua facit per decem mille cilias suas. Signum "ciliarum" paramecii quoque habent nonnulla unicellularia alia qui etiam in phylo Ciliophororum numerantur. Obstacula et angusta perlabi potest faciliter, quia cutem elasticam habet. Hostes sunt amoebae et protistum ciliatum quoddam (Didinium nasutum). Paramecia habent Trichocystas ad se ipsa defendenda filameta proteinosa ac glutinosa eiciendis.
pagina theodisca Pantoffeltierchen
Klumpelės (lot. Paramecium) – vienaląsčių pirmuonių gentis, judanti blakstienėlėmis, paplitusi visame pasaulyje.
Klumpelės turi du branduolius, iš kurių vienas dalyvauja dauginantis, o kitas naudojamas „ląstelės ūkiui“ tvarkyti. Tai gana dideli (0,05-0,3 mm) pirmuonys, juos pasistengus galima matyti net ir plika akimi.
Rūšis:
Paramesium ialah protozoa air tawar dalam genus PARAMESIUM, badannya bujur, diliputi membran plasma yang berbulu (silia), berbentuk tetap seperti selipar yang leper dan mempunyai alur berbentuk corong untuk makan (mulut).
Paramesium tersebar luas di persekitaran air tawar, dan terutamanya biasa dalam kumbahan. Kini beberapa spesies baru Paramesium telah dijumpai di lautan.
Sesetengah eukaryotes sel tunggal, seperti Paramecium, merupakan contoh pengecualian kepada kod genetik sejagat: dalam sistem terjemahan mereka beberapa codon berbeza dari genetik piwaian.
Sel menyerupai prolate spheroid,[1] bulat di hadapan dan runcing di belakang. Pellicle adalah kaku tetapi selaput lentur memberikan Paramecium bentuk khususnya. Menyelitupi bahagian luarannya adalah struktur seperti rerambut, dikenali sebagai silia. Pada bahagian sisi, bermula di hadapan dan berterusan separuh jalan, adalah liang oral, yang mengumpulkan makanan sehingga ia di sapu masuk ke mulut sel. Terdapat bukaan pada bahagian belakang yang dikenali sebagai liang dubur ("anal pore"). Vakuol contractile dan salurannya yang tersebar, dirujuk sebelumnya sebagai pengosmokawalaturan organisma, juga terdapat di luar Paramesium. Paramesium biasanya dikelirukan dengan Blepharisma.
Paramecium mengandungi sitoplasma, trichocyst, tekak, vakuol makanan, makronukleus, dan mikronukleus. Ia juga memiliki vakuol mengecut yang menyingkirkan air agar sel tidak pecah kerana air yang berlebihan yang bebas memasuki selaput disebabkan oleh sifat separa telap selaput.
Silia merupakan struktur pergerakan lokomotif Paramecium. Agar Paramecium boleh bergerak ke hadapan, silianya bergerak menurut rentak pada sudut 120 darjah, ke belakang secara serentak (iaitu, cilium mengeliat dari hujung ke pangkal). Ini bererti bahawa Paramecium bergerak dengan sulur melalui air di paksi yang maya. Paramecium juga boleh bergerak ke belakang apabila silia mengepuk ke hadapan pada sudut dengan serentak.
Sekiranya Paramecium melanggar halangan ia mengundur, belok sedikit dan bergerak ke hadapan lagi. Jika ia kembali melanggar objek pepejal sekali lagi, ia akan mengulangi proses ini sehingga ia melepasi objek tersebut.
Paramecia makan organisma mikro seperti bakteria, alga, dan yis. Untuk mengumpulkan makanannya, Paramecium menggunakan cilianya bagi menyapu makanannya sepanjang bersama air ke dalam mulut sel selepas ia jatuh ke dalam liang tekak ("oral groove"). Makanan masuk melalui mulut sel kedalam perutnya. Apabila makanan yang mencukupi telah terkumpul di pangkal tekak, ia membentuk satu vakuol makanan dalam sitoplasma, dan bergerak melalui sel itu, melalui bahagian belakang terlebih dahulu. Ketika ia bergerak, enzim daripada sitoplasma memasuki vakuol untuk mencerna kandungan, nutrien yang dicerna kemudian memasuki sitoplasma, dan vakuol mengecut. Apabila vakuol sampai ke liang dubur, ia pecah, menyingkirkan sisa kandungan kepada bahagian luar.
Salah satu hubungan simbiotik yang paling menarik yang diketahui ialah Paramecium aurelia dan bakteria endosymbionts. Lihat juga Chlorella simbiosis dengan Paramecium bursaria.
Genom Paramecium telah dijujukkan (spesies: Paramecium tetraurelia), memberi bukti bagi salinan tiga pertindihan keseluruhan genom.[2]
Bagi sesetengah ciliates, seperti Stylonychia dan Paramecium, hanya UGA dinyahkod sebagai codon henti, sementara UAG dan UAA diatur kembali sebagai codon deria.[3]
Persoalan mengenai samaada paramecia memaparkan pembelajaran telah menjadi objek banyak ujikaji, menghasilkan keputusan yang bercampur-campur. Dalam salah satu ujikaji paling terkini yang diterbitkan,[4] pengarang, dengan menggunakan voltan sebagai penggalak, menyimpulkan bahawa Paramecium mampu membezakan antara tahap keterangan yang berbeza.
|month= tidak diketahui diabaikan (bantuan)Selenggaraan CS1: Penggunaan eksplisit et al. (link) Selenggaraan CS1: Pelbagai nama: senarai pengarang (link) |month= tidak diketahui diabaikan (bantuan)Selenggaraan CS1: Pelbagai nama: senarai pengarang (link)
Paramesium ialah protozoa air tawar dalam genus PARAMESIUM, badannya bujur, diliputi membran plasma yang berbulu (silia), berbentuk tetap seperti selipar yang leper dan mempunyai alur berbentuk corong untuk makan (mulut).
Paramesium tersebar luas di persekitaran air tawar, dan terutamanya biasa dalam kumbahan. Kini beberapa spesies baru Paramesium telah dijumpai di lautan.
Sesetengah eukaryotes sel tunggal, seperti Paramecium, merupakan contoh pengecualian kepada kod genetik sejagat: dalam sistem terjemahan mereka beberapa codon berbeza dari genetik piwaian.
Pantoffeldiertjes (wetenschappelijke naam: Paramecium) is een geslacht van eencellige organismen uit de stam van de Ciliophora (trilhaardiertjes).
Afhankelijk van de soort bedraagt de lengte tussen de 50 en 300 micrometer.
De vorm van de cel lijkt enigszins op een klein pantoffeltje. Het lichaamsoppervlak is bedekt met trilharen die zorgen voor de voortbeweging. Door de asymmetrische vorm draait het diertje zich om zijn lengteas rond als het zich met behulp van de trilharen verplaatst. De trilharen hebben ook een functie bij het voeden.
Een pantoffeldiertje bevat twee typen kernen, één macronucleus die betrokken is bij de metabolische functies van de cel, en één micronucleus die een rol speelt bij de voortplanting.
Pantoffeldiertjes zijn bacterivoor: ze voeden zich met van afgestorven plantenresten levende bacteriën. Het milieu waar de pantoffeldiertjes leven is zuurstofarm tot praktisch zuurstofloos door het rottingsproces dat door de bacteriën veroorzaakt wordt.
Om voedsel op te nemen brengt het pantoffeldiertje de voedseldeeltjes met de cilia naar de orale groeve (peristoom). In deze orale groeve ligt het cytostoom, een voorafbepaalde plaats waar het pantoffeldiertje via instulping een voedselvacuole maakt rond de voedseldeeltjes. Deze voedselvacuolen maken dan een reis door het lichaam, waarbij het voedsel wordt verteerd. Het onverteerde materiaal wordt uitgescheiden aan het cytoproct.
Het pantoffeldiertje heeft twee alternerend kloppende vacuoles. Er is een constante osmotische beweging van water van buiten naar binnen het lichaam. Via de kloppende vacuolen wordt het overtollige water weggepompt, zodat het pantoffeldiertje niet "ontploft". Als een kloppende vacuole samentrekt verlaat het water via een porie het lichaam. Die porie wordt afgebroken als de vacuole leeg is en wordt weer opgebouwd als de vacuole vol is. Dit geldt ook voor de verzamelkanalen waarlangs de vacuole het water verzamelt. Deze worden afgebroken als de vacuole vol is (zodat het water tijdens de pompbeweging niet langs die kanalen wegloopt) en worden weer opgebouwd als de vacuole leeg is (zodat nieuw water verzameld kan worden).
Zelf zijn pantoffeldiertjes het voedsel van vele kleine vissen en andere waterdiertjes.
Paramecium bursaria is de gastheer van een endosymbiont, de groenalg Chlorella, waarvan er per gastheer honderden aanwezig kunnen zijn. Deze endosymbionten hebben elk een eigen vacuole. Ze voorzien de gastheer van suikers en maken fotoautotrofe levenswijze mogelijk. De symbiose is facultatief: beide partners kunnen onafhankelijk van elkaar groeien, maar de gastheer kan de endosymbiont weer opnemen, nadat deze verloren is gegaan, bijvoorbeeld doordat de endosymbiont door de gastheer is verteerd tijdens het leven in het donker.
De voortplanting is meestal ongeslachtelijk door celdeling. De geslachtelijke voortplanting vindt plaats tijdens een bepaald deel van de levenscyclus. Dit proces zorgt voor genetische uitwisseling door zowel het laten plaatsvinden van meiotische als mitotische celdelingen. Deze vorm van voortplanting wordt ook wel conjugatie genoemd. Als de (over)leefomstandigheden goed genoeg zijn, zal de voortplanting voornamelijk plaatsvinden door mitotische celdelingen. Het is de snelste en makkelijkste manier van voortplanting en er zijn geen aanpassingen van het DNA nodig om te kunnen overleven. Als de (over)leefomstandigheden niet goed zijn, zal de voortplanting voornamelijk plaatsvinden door meiotische celdelingen. Om kansen voor een beter DNA voor in deze omgeving met de negatieve leefomstandigheden, zodat ook deze leefomgeving bewoond kan worden. DNA wordt tussen twee organismen uitgewisseld door meiose die plaats heeft gevonden in de micronucleus wat uitgewisseld wordt.
De volgende soorten worden te onderscheiden:[1]
Pantoffeldiertjes (wetenschappelijke naam: Paramecium) is een geslacht van eencellige organismen uit de stam van de Ciliophora (trilhaardiertjes).
Tøffeldyr (Paramecium) er ei gruppe eincella organismar. Dei har namnet frå si noko tøffel-aktige form. Dei vert 50-350 mikrometer lange og er kledd med flimmerhår. Dei sym ved å laga bølgjande rørsler med flimmerhåra. Viss dei vert forstyrra, snur dei symjerørsla.[1] Dei er vanlege i stillestående vatn. Tøffeldyr lever stort sett av bakteriar.[1]
Tøffeldyr (Paramecium) er ei gruppe eincella organismar. Dei har namnet frå si noko tøffel-aktige form. Dei vert 50-350 mikrometer lange og er kledd med flimmerhår. Dei sym ved å laga bølgjande rørsler med flimmerhåra. Viss dei vert forstyrra, snur dei symjerørsla. Dei er vanlege i stillestående vatn. Tøffeldyr lever stort sett av bakteriar.
Tøffeldyr er en gruppe encellede organismer. De har navnet fra sin noe tøffel-aktige form. De blir 50-350 mikrometer lange og er kledd med flimmerhår. De svømmer ved å lage bølgende bevegelser med flimmerhårene. Hvis de blir forstyrret, snur de svømmebevegelsen.[1] De er vanlige i stillestående vann. De lever stort sett av bakterier og andre små celler.
Tøffeldyr er en gruppe encellede organismer. De har navnet fra sin noe tøffel-aktige form. De blir 50-350 mikrometer lange og er kledd med flimmerhår. De svømmer ved å lage bølgende bevegelser med flimmerhårene. Hvis de blir forstyrret, snur de svømmebevegelsen. De er vanlige i stillestående vann. De lever stort sett av bakterier og andre små celler.

Pantofelki (Paramecium) – rodzaj „protistów zwierzęcych” (Pierwotniaki) należące do typu orzęsków (Ciliata) z podgromady równorzęsych.
Protisty należące do tego rodzaju mają kształt wydłużony, cygarowaty, owalny lub pantofelkowaty. Osiągają rozmiar od 120 μm do 350 μm. Ciało z zewnątrz pokryte jest rzędami rzęsek oraz wyposażone w tzw. trichocysty. Rzęski pokrywające ciało służą do poruszania się i zdobywania pokarmu. Ciało otacza żywa osłona zwana pellikulą.
Systematyka tego rodzaju zmienia się w dalszym ciągu. Status części gatunków jest niepewny[1]. Do rodzaju tego należą następujące gatunki pantofelków[2]:
Pantofelki (Paramecium) – rodzaj „protistów zwierzęcych” (Pierwotniaki) należące do typu orzęsków (Ciliata) z podgromady równorzęsych.
Protisty należące do tego rodzaju mają kształt wydłużony, cygarowaty, owalny lub pantofelkowaty. Osiągają rozmiar od 120 μm do 350 μm. Ciało z zewnątrz pokryte jest rzędami rzęsek oraz wyposażone w tzw. trichocysty. Rzęski pokrywające ciało służą do poruszania się i zdobywania pokarmu. Ciało otacza żywa osłona zwana pellikulą.
Systematyka tego rodzaju zmienia się w dalszym ciągu. Status części gatunków jest niepewny. Do rodzaju tego należą następujące gatunki pantofelków:
Paramecium aurelia Paramecium biaurelia Paramecium bursaria Paramecium calkinsi (Woodruff, 1921) Pantofelek (Paramecium caudatum) Paramecium decaurelia Paramecium dodecaurelia Paramecium duboscqui Paramecium jenningsi Paramecium multimicronucleatum Paramecium nephridiatum Paramecium novaurelia Paramecium octaurelia Paramecium pentaurelia Paramecium polycarum Paramecium polycaryum Paramecium primaurelia Paramecium putrinum Paramecium quadecaurelia Paramecium schewiakoffi Paramecium septaurelia Paramecium sexaurelia Paramecium sonneborni Paramecium tetraurelia Paramecium tredecaurelia Paramecium triaurelia Paramecium undecaurelia Paramecium woodruffiParamecium é um género bem conhecido de protozoários ciliados,[1] com dimensões entre 50 e 300 micrômetros, dependendo das espécies. O corpo das paramécias é coberto de cílios simples, mas o sulco oral (a "boca") contém pequenos cílios orais compostos, que são típicos da ordem Peniculida. As paramécias habitam em ambientes de água doce, salobra e marinha, e também são facilmente encontradas em poças d'água e em locais com água parada. Pode ingerir cerca de 5.000.000 leveduras em 24 horas.
O Paramecium, como protozoário cilíado tem características peculiares a serem definidas. Por exemplo, além de os cílios ajudarem na sua locomoção, são também usados para trazer alimentos para dentro do citóstoma onde serão fagocitados após passar pela citofaringe.
Os organismos citoplasmáticos contituintes do Paramecium são:
O paramecium tem reprodução sexuada realizada por meio de conjugação, que se dá em etapas.
Primeiro os paramécios ligam-se pelos citóstomas.
O macronúcleo dos dois paramécios degeneram, e os seus micronúcleos(2n) por meio de meiose geram quatro novos micronúcleos haploides.
Desses oito novos micronúcleos formados, seis se degeneram, sobrando apenas um em cada paramécio. Logo esses que sobraram sofrem mitose e geram dois, que são trocados pelos paramécios entre si por meio do citóstoma.
Dentro do paramécio esses micronúcleos juntam-se formando a ploidia 2n novamente. Os paramécios separam-se.
Depois disso, os micronúcleos sofrem duas mitoses sucessivas, que geram quatro micronúcleos diploides. Dois desses desenvolvem para Macronúcleos, e dois continuam sendo o que eram. A célula sofre bipartição gerando dois paramécios e finalizando o processo sexuado.
O processo assexuado é simples. Ocorre por meio de cissiparidade ou bipartição, ou seja o paramécio duplica seus núcleos e se divide, gerando cópias de si.
A reprodução por divisão leva cerca de duas horas.
O Paramecium é um himenostomado. É um livre-natante de águas estagnadas (encontrado em água doce e fundos lodosos onde há matéria orgânica em decomposição), de movimentos rápidos por causa dos cílios. A corrente continua de água que os cílios provocam também se envolve na sua alimentação, que se compõe de algas e bactérias microscópicas, ou, algumas poucas gotas de leite por dia.
O género Paramecium foi descrito pela primeira vez no século XVIII, mas a divisão em espécies ainda não está completamente esclarecida - por exemplo, P. aurelia foi recentemente dividida em 14 espécies. A lista seguinte contém espécies geralmente reconhecidas:
São criados em recipientes de vidros ou garrafas de 2 L, com certa facilidade, em culturas com folhas de alface ou de couve, secas ao sol, em infusão na água, se possível estagnada. Vive em média 13 dias. São o alimento inicial ideal para alevinos.[2]
Paramecium é um género bem conhecido de protozoários ciliados, com dimensões entre 50 e 300 micrômetros, dependendo das espécies. O corpo das paramécias é coberto de cílios simples, mas o sulco oral (a "boca") contém pequenos cílios orais compostos, que são típicos da ordem Peniculida. As paramécias habitam em ambientes de água doce, salobra e marinha, e também são facilmente encontradas em poças d'água e em locais com água parada. Pode ingerir cerca de 5.000.000 leveduras em 24 horas.
Parameciul (Paramecium) reprezintă cel mai evoluat gen dintre protistele animaloide. Cuprinde mai multe specii: Paramecium caudatum, Paramecium aurelia, Paramecium tetraurelia. Sunt unicelulare, cu lungimea de aproximativ 0,25 mm sau 50 până la 350 µm.
Corp alungit, de forma unui „pantofior”, acoperit cu numeroși cili scurți și uniformi. Pe fața ventrală a corpului prezintă o depresiune oblică numită peristom, în capătul peristomului se află citostomul, care se continuă cu un citofaringe, la capătul acestuia desprinzându-se vacuole digestive. La interior conține organite celulare, două vacuole contractile (cu rol excretor și osmoreglator) și doi nuclei (un macronucleu cu rol metabolic și un micronucleu cu rol genetic).
Se face prin toată suprafața corpului.
Paramecii au un mod de nutriție holozoic, pot fi considerate răpitoare. Cavitatea bucală este numită peristom, care se prelungește în interior cu un citostom și se termină cu un citofaringe. Hrana, pătrunzând în interior prin aceste structuri, este înglobată la capătul citofaringelui într-o vacuolă digestivă care este transportată prin întregul microorganism cu ajutorul curentului citoplasmatic, în interiorul acestor vacuole sub acțiunea unor enzime are loc digestia hranei, iar substanțele nutritive sunt transportate direct la organisme pentru realizarea celorlalte procese vitale.
Are loc cu ajutorul vacuolelor contractile, care funcționează ritmic. La început, vacuola crește în talie, până atinge un volum maxim, apoi ea se contractă rapid, eliminând lichidul conținut în mediul ambiant. Vacuola contractilă poate lipsi la unele forme marine sau parazite, însă prezența ei la formele dulcicole este obligatorie. Vacuola contractilă este înconjurată de un sistem de canalicule colectoare, aceste canale la rândul lor sunt înconjurare de o citoplasmă excretoare numită spongiom .
La parameci, reproducerea se realizează atât pe cale asexuată, cât și sexuată. Asexuat prin diviziune transversală și în condiții nefavorabile ale mediului se pot divide sub un chist. Sexuat prin conjugare [1] . La Paramecium caudatum conjugarea are loc în felul următor: 2 indivizi se alipesc prin intermediul peristoamelor, creându-se între aceștia o punte citoplasmatică. Având în vedere faptul că sunt prezenți 2 nuclei în interiorul fiecăruia dintre conjugați, macronucleu și micronucleu, primul dintre nuclei nu participă la cunjugare și astfel macronucleul se resoarbe, iar micronucleul suferă două diviziuni succesive, care duc la formarea în total a 4 nuclei, la rândul lor 3 dintre nuclei se resorb, iar al 4-lea se mai divide odată, dând naștere la 2 nuclei haploizi. Unul din nuclei este staționar (considerat femel), iar celălalt este „migrator” (considerat mascul). Procesul descris are loc în ambii parameci. În faza următoare are loc schimbul de nuclei migranți prin intermediul punții citoplasmatice și fuzionarea lor cu nucleii staționari, rezultându-se „sincarionul” care are valoare de zigot diploid. Imediat după acesta, paramecii se separă devenind exconjugați. În fiecare exconjugat, sincarionul suferă trei diviziuni succesive și are loc formarea a 8 nuclei, 4 dintre aceștia evoluează în macronucleu, unul în micronucleu, iar 3 se resorb. În continuare parameciul se divide de două ori, rezultând din fiecare exconjugat câte 4 indivizi noi.
Prin modul de hrănire asigură purificarea apelor
Parameciul (Paramecium) reprezintă cel mai evoluat gen dintre protistele animaloide. Cuprinde mai multe specii: Paramecium caudatum, Paramecium aurelia, Paramecium tetraurelia. Sunt unicelulare, cu lungimea de aproximativ 0,25 mm sau 50 până la 350 µm.
Paramecium tetraurelia
Paramecium aurelia
Paramecium caudatum
Paramecij je pražival iz rodu migetalkarjev. V dolžino meri 50-350 μm, odvisno od vrste.
Zanj sta značilni dve jedri v citoplazmi; eno je namenjeno razmnoževanju, drugo pa življenjskim potrebam. Hrani se z bakterijami in drugimi mikroorganizmi. Razmnožuje se nespolno z delitvijo ali pa spolno s konjugacijo. Paramecij je široko razširjen v sladkih in morskih vodah.
Paramecij je pražival iz rodu migetalkarjev. V dolžino meri 50-350 μm, odvisno od vrste.
Zanj sta značilni dve jedri v citoplazmi; eno je namenjeno razmnoževanju, drugo pa življenjskim potrebam. Hrani se z bakterijami in drugimi mikroorganizmi. Razmnožuje se nespolno z delitvijo ali pa spolno s konjugacijo. Paramecij je široko razširjen v sladkih in morskih vodah.
Toffeldjur (Paramecium) är ett släkte av encelliga organismer som ingår i stammen ciliater. Toffeldjuren är cirka 50 mikrometer breda och 150 mikrometer långa och klädda med flimmerhår (cilier).[1] De rör sig genom böljande rörelser från flimmerhåren och simmar på det viset. Toffeldjur lever främst i sötvatten och är mycket vanliga i stillastående vatten. De flesta är heterotrofer och lever på bakterier och andra mikroorganismer men en del är mixotrofer och får en del av sin näring från symbios med encelliga alger vilket gör dess förmåga att anpassa sig bättre.[2] Ett toffeldjur förökar sig genom delning.[förtydliga] Toffeldjur är lätta att föröka i laboratorium och används därför ofta som modellorganismer inom genetik och cellbiologi.[1]
Toffeldjur (Paramecium) är ett släkte av encelliga organismer som ingår i stammen ciliater. Toffeldjuren är cirka 50 mikrometer breda och 150 mikrometer långa och klädda med flimmerhår (cilier). De rör sig genom böljande rörelser från flimmerhåren och simmar på det viset. Toffeldjur lever främst i sötvatten och är mycket vanliga i stillastående vatten. De flesta är heterotrofer och lever på bakterier och andra mikroorganismer men en del är mixotrofer och får en del av sin näring från symbios med encelliga alger vilket gör dess förmåga att anpassa sig bättre. Ett toffeldjur förökar sig genom delning.[förtydliga] Toffeldjur är lätta att föröka i laboratorium och används därför ofta som modellorganismer inom genetik och cellbiologi.
Terliksi hayvan, paramesyumlar (Paramecium) olarak da bilinir, silli protozoaların en çok bilinen örneklerinden birisidir; genellikle silli protozoaların temsilcileri olarak çalışılır. Terliksi hayvan; tek hücreli ve terlik şeklindedirler ("terliksi hayvan" ismini alma sebepleri budur). Boyları, türlerine göre 30 veya 500 mikrometre arasında değişir. Basit silleri, vücutlarını kaplar; ve diğer peniculidlerde olduğu gibi göze çarpmayacak şekilde birleşik ağızsal siller de içeren derin bir ağızsal boşlukları bulunur.
Ozmoregülasyon işlemi, etraftan ozmos basınçla emilmiş olan suyu aktif olarak dışarı çıkaran bir çift kontraktif koful tarafından gerçekleştirilir. Terliksi hayvanlara genellikle temiz su ortamlarında ve kir tabakalarında rastlamak mümkündür. Terliksi hayvanlar, asidik ortamlara ilgi gösterirler. Genellikle aseksüel olarak ürerler; ama yeterli besin maddelerinin bulunmadığı ortamlarda da konjugasyona başvurabilirler.
Terliksi hayvanların türlere ayrımında şimdilik hala bazı değişikler yapılmaktadır; örneğin P. aurelia kısa bir süre önce 14 türe ayrılmıştır; geriye kalanlar sırasıyla:
Terliksi hayvan, paramesyumlar (Paramecium) olarak da bilinir, silli protozoaların en çok bilinen örneklerinden birisidir; genellikle silli protozoaların temsilcileri olarak çalışılır. Terliksi hayvan; tek hücreli ve terlik şeklindedirler ("terliksi hayvan" ismini alma sebepleri budur). Boyları, türlerine göre 30 veya 500 mikrometre arasında değişir. Basit silleri, vücutlarını kaplar; ve diğer peniculidlerde olduğu gibi göze çarpmayacak şekilde birleşik ağızsal siller de içeren derin bir ağızsal boşlukları bulunur.
Ozmoregülasyon işlemi, etraftan ozmos basınçla emilmiş olan suyu aktif olarak dışarı çıkaran bir çift kontraktif koful tarafından gerçekleştirilir. Terliksi hayvanlara genellikle temiz su ortamlarında ve kir tabakalarında rastlamak mümkündür. Terliksi hayvanlar, asidik ortamlara ilgi gösterirler. Genellikle aseksüel olarak ürerler; ama yeterli besin maddelerinin bulunmadığı ortamlarda da konjugasyona başvurabilirler.
Terliksi hayvanların türlere ayrımında şimdilik hala bazı değişikler yapılmaktadır; örneğin P. aurelia kısa bir süre önce 14 türe ayrılmıştır; geriye kalanlar sırasıyla:
Paramecium aurelia Ehrenberg, 1838 Paramecium bursaria (Ehrenberg) Focker, 1836 P. calkinsi Woodruff, 1921 P. caudatum Ehrenberg, 1838 P. duboscqui Chatton and Brachon, 1933 P. jenningsi Diller & Earl, 1958 P. multimicronucleatum Powers & Mitchell, 1910 P. nephridiatum von Gelei, 1925 P. polycaryum Woodruff, 1923 P. putrinum Claparede & Lachmann, 1858 P. trichium Stokes, 1885 P. woodruffi Wenrich, 1928Війки вкривають тіло парамецій, їх скорочення дозволяє клітині рухатися вперед. Ротова борозенка покрита навколоротовими війками, які приганяють їжу в клітинну глотку парамецій. Живляться бактеріями й іншими дрібними одноклітинними організмами. Деякі види (Paramecium bursaria) мають ендосимбіонтів-зоохлорел, від яких отримують більшу частину їжі. Відомий також симбіоз Paramecium aurelia з бактеріями (каппа-частками), який перетворює штами інфузорій в «кілерні». Кілерні штами виділяють в навколишнє середовище каппа-частинки, які при поглинанні їх чутливими (позбавленими каппа-частинок) штамами викликає їх загибель. Осморегуляція здійснюється двома скоротливими вакуолями, які активно виводять з клітки воду, що поступила з навколишнього середовища. Парамеціі віддають перевагу кислим умовам середовища.
Trùng đế giày (còn gọi là Paramecium, trùng cỏ, trùng giày hay thảo trùng) là đại diện của lớp Trùng cỏ. Tế bào trùng đế giày đã phân hóa thành nhiều bộ phận. Mỗi bộ phận đảm nhận một chức năng sống nhất định.
Trùng giày được con người biết đến trước tiên trong thế giới động vật đơn bào. Khi chế tạo được kính hiển vi, người ta thử lấy nước "cỏ ngậm" soi thì tình cờ phát hiện ra chúng và vì thế được gọi là "trùng cỏ". Ngày nay "trùng cỏ" trở thành tên chính thức cho nhóm động vật này và nước "cỏ ngậm" vẫn là môi trường nuôi cấy trùng cỏ lý tưởng ở phòng thí nghiệm.
Có hình giống đế giày.[1] Cơ thể có hình khối, không đối xứng, giống chiếc giày. Trùng di chuyển nhờ lông bơi. Phần giữa cơ thể là bộ nhân gồm: nhân lớn và nhân nhỏ. Nửa trước và nửa sau đều có 1 không bào co bóp hình hoa thị và ở 1 vị trí cố định. Chỗ lõm của cơ thể là rãnh miệng, cuối rãnh miệng có lỗ miệng và hầu. Mỗi bộ phận đảm nhiệm chức năng sống nhất định.
Trùng giày là động vật đơn bào đã có sự phân hóa thành các bộ phận như nhân lớn, nhân nhỏ, không bào co bóp. Thức ăn (gồm vi khuẩn, vụn hữu cơ,...) được lông bơi dồn về lỗ miệng. Thức ăn qua miệng và hầu được vo thành viên trong không bào tiêu hóa. Sau đó không bào tiêu hóa rời hầu di chuyển trong cơ thể theo một quỹ đạo nhất định, Enzim tiêu hóa biến thức ăn thành chất lỏng thấm vào chất nguyên sinh nuôi cơ thể. Chất bã được thải ra ngoài lỗ thoát ở thành cơ thể.
Ngoài hình thức sinh sản vô tính bằng cách phân đôi theo chiều ngang, trùng giày còn có hình thức sinh sản hữu tính gọi là sinh sản tiếp hợp. Chúng phân đôi trung bình gần 1 lần mỗi ngày. Quá trình tiếp hợp xảy ra có thể kéo dài tới 12 giờ ở điều kiện 16 độ C.
Trùng giày sống ở những váng cống rãnh hoặc những váng nước đục. Mỗi khi đời sống gặp khó khăn (khô hạn hay thiếu thức ăn), trùng đế giày có khả năng tiết nước thừa, thu nhỏ cơ thể lại, tiết ra lớp vỏ bọc gọi là hóa bào xác. Trong bào xác, chúng tồn tại rất lâu và có cơ hội được gió cuốn đi để phát tán đến những môi trường mới thích hợp hơn trước..
Trùng giày bơi rất nhanh trong nước nhờ lông bơi, vừa tiến vừa xoay.
Trùng giày có ích lợi trong việc đóng vai trò là một mắt xích trong chuỗi thức ăn của tự nhiên.


undefined
Trùng đế giày (còn gọi là Paramecium, trùng cỏ, trùng giày hay thảo trùng) là đại diện của lớp Trùng cỏ. Tế bào trùng đế giày đã phân hóa thành nhiều bộ phận. Mỗi bộ phận đảm nhận một chức năng sống nhất định.
Trùng giày được con người biết đến trước tiên trong thế giới động vật đơn bào. Khi chế tạo được kính hiển vi, người ta thử lấy nước "cỏ ngậm" soi thì tình cờ phát hiện ra chúng và vì thế được gọi là "trùng cỏ". Ngày nay "trùng cỏ" trở thành tên chính thức cho nhóm động vật này và nước "cỏ ngậm" vẫn là môi trường nuôi cấy trùng cỏ lý tưởng ở phòng thí nghiệm.
Параме́ции, или инфузории туфельки[1] (лат. Paramecium) — род инфузорий, включающий несколько сотен видов, в том числе множество видов-двойников. Длина тела различных представителей составляет от 50 до 350 микрометров.
Реснички покрывают тело простейшего, их биение позволяет клетке двигаться вперёд. Ротовая бороздка покрыта околоротовыми ресничками, которые пригоняют пищу в клеточную глотку парамеций. Питаются парамеции бактериями и другими мелкими одноклеточными организмами. Некоторые виды (Paramecium bursaria) имеют эндосимбионтов-зоохлорелл, от которых получают бо́льшую часть пищи. Известен также симбиоз Paramecium aurelia с бактериями (каппа-частицами), который превращает штаммы инфузорий в «киллерные». Киллерные штаммы выделяют в среду каппа-частицы, которые при поглощении их чувствительными (лишёнными каппа-частиц) штаммами вызывает их гибель. Осморегуляция осуществляется двумя сократительными вакуолями, которые активно выводят из клетки воду, поступившую из окружающей среды. Парамеции предпочитают кислые условия среды.
Геном одного из видов (Paramecium tetraurelia) полностью расшифрован. Как было установлено, он содержит около 40 000 кодирующих белки генов (примерно в два раза больше, чем человеческий). Показано, что число генов увеличилось в результате трёх последовательных дупликаций исходного генома, последняя из которых произошла в период возникновения видов-двойников комплекса P. teraurelia[2]. Генетический код у парамеций отличается от универсального генетического кода, в частности, у них есть только один кодон, обозначающий конец синтеза полипетидной цепи (стоп-кодон) (обычно их три)[3].
Paramecium bursaria, вид вместе с симбиотической водорослью
Paramecium putrinum
Paramecium
Конъюгация Paramecium caudatum
Параме́ции, или инфузории туфельки (лат. Paramecium) — род инфузорий, включающий несколько сотен видов, в том числе множество видов-двойников. Длина тела различных представителей составляет от 50 до 350 микрометров.
Paramecium aurelia
Paramecium bursaria
Paramecium caudatum
Paramecium tetraurelia
多數古老的單細胞生物,形狀都不是很固定,有的甚至無法辨認其外形。然而草履蟲就很容易辨別,其身体呈圆筒形,前端较圆,中后部较宽,后端较尖,平面看形状像倒置的草鞋,故名。
身長約零點一至零點三毫米,常见的是大草履虫(p.caudatum),生活在有淡水的地方,肉眼勉強可見模糊的外形,需用複式顯微鏡觀察才能看清楚。在顯微鏡下,可觀察到牠整個構造就是一個細胞,是一種單細胞生物,在這個細胞體內含有兩個細胞核、兩個伸縮泡和一些食物泡,全身纵行排列布满大致同长的细纤毛。
草履蟲是利用身上一種排列方式特殊的維管負責纖毛的擺動得以前進,是為原生動物中運動速度最快的一類,當草履蟲游泳時,纖毛的擺動會如同船槳一般,一收一放,藉此得以推動身體前進。若於前進同時,遇到了障礙物,纖毛就會產生相反方向的運動,先後退再換方向前進。
草履蟲主要以水中细菌及其他水裡的有机物作为食物,对不同的细菌有选食性。草履蟲的纖毛擺動不但能幫助牠移動身軀,同時也可藉此擺動使食物隨著水流送入口溝,形成食物泡,然後食物随着细胞质流动在泡内逐步消化,不能消化的残渣再通过表膜上开口的細胞肛孔排出体外。
輪蟲或其他原生動物是牠主要的天敵。草履蟲主要的武器是絲細胞,其在表膜下整齐埋藏许多刺丝泡,当受到强烈刺激(化学、电、机械等)或敌害时,在2~3微秒内可射出刺丝泡,变成一根长约40微米的细丝,前端有矛状小针,把毒素做絲狀噴出;有的草履蟲其絲細胞不含毒素,但可以噴出絲狀物,用來纏繞敵人或嚇走對方,更可用這種絲細胞來捕捉食物。草履虫靠体纤毛的摆动在水中游动,身体反时针方向旋转前进。遇陣碍物或微弱刺激时,纤毛朝相反方向击打,改变游动路线以避开。

ゾウリムシは、顕微鏡下では草履(ぞうり)のような形に見える繊毛虫の1種 Paramecium caudatum の和名、広義にはゾウリムシ属 (Paramecium) に属する種を指す。単細胞生物としてはよく名を知られている。微生物自体の発見者であるオランダのレーウェンフックによって17世紀末に発見された。日本語名は、動物学者の川村多実二が1930年につけたものであり、英語名の「slipper animalcule」の「slipper」を「草履」と意訳したことに由来している。
分類表内の写真は、Paramecium aurelia の位相差顕微鏡像である。色は位相差の光学系に依るものであり、細胞本来の色ではない。楕円形の細胞周囲を取り囲み薄紫色に写っている部分が移動に用いる繊毛である。右下と右上に白くわずかに星型の輪郭が見える目立つ構造は浸透圧を調整するための収縮胞。中央やや下に見えるのっぺりした灰色の部分が栄養核として知られる大核。多数の食胞も見える。細胞口、細胞肛門、生殖用の小核などはこの写真では見えない。
細胞の長さは 90-150µm、幅は 40µm 程度である。名前は平たい印象を与えるが実際には円筒形に近く、中腹には細胞口というくぼみがややねじれるように入っている。細胞表面には約3500本の繊毛を持っており、その繊毛を使って泳ぐ。繊毛は体表の繊毛列にそって生えている。繊毛は細胞全体にほぼ均一に生えているが、細胞口の奥の部分では細胞咽頭に向けて特殊な配置と動きが見られる。繊毛は5%エタノール処理などのストレスにより脱離させることができる(脱繊毛)。細胞口の奥には細胞内へ餌を取り込む細胞咽口があり、餌はここを通って食胞に取り込まれる。食胞内で消化が行われ、有用な成分は細胞内へ吸収されながら、食胞は細胞内を回るように移動する。排泄物は細胞後方の細胞肛門から放出される。
細胞の前後には、大きな星形もしくは花に見える放射状の細胞器官がある。これを収縮胞と言い、細胞内の浸透圧調節を担っている。収縮胞は中央の円形の部分と、周囲に花びらのように並ぶ涙滴型の部分から構成される。水の排出時にはまず涙滴型の部分に水が集まり、ここから中央の円形の部分に水が移され、細胞外に水が放出される。
細胞内には大小2つの細胞核があり、それぞれ大核と小核と呼ばれる。大核は普段の活動に関わる。小核は生殖核とも呼ばれ、有性生殖に関して働くとされる。細胞内に機能的に分化した核を持つのは繊毛虫類の特徴である。
無性生殖は分裂による。他の繊毛虫同様、体軸方向の前後の部分に分かれるようにして分裂する。有性生殖としては細胞の接合が行われるが、その方法はやや特殊である。接合に先立ち、大核が消失するとともに生殖核である小核が減数分裂を行い、4つの核に分かれる。このうち3つは消失し、残った1つがさらに2つに分裂し、このうち1つの核を互いに交換する。その後、それぞれの細胞内の2核が融合することで接合は完了する。この間、2個体のゾウリムシは互いに同一方向を向いて寄り添うが、細胞間に連絡を持つだけで細胞そのものの融合は行われない。なお、接合後に大核は小核を元に改めて形成される。
水田や沼や池など、淡水の止水域に分布する。細胞表面の繊毛により遊泳するため、単細胞生物としては移動力が大きい。障害物などに接触すると、繊毛逆転により遊泳方向を反転する(後退遊泳)。
ゾウリムシは主に真正細菌を餌とする。ただし、近似種のミドリゾウリムシ(Paramecium bursaria)は体内に緑藻であるクロレラを共生させており、光合成産物の還流を受けて生活することも可能である。逆に、ゾウリムシの捕食者は大型のアメーバや、ディディニウム(Didinium、シオカメウズムシ)といった他の繊毛虫である。ディディニウムは細胞前端の口吻部にエクストルソーム(extrusome)と呼ばれる射出器官を持ち、これをゾウリムシに打ち込んで動きを止め、飲み込んで消化する。
ゾウリムシは浸透栄養ではなく捕食性であるため、培養に際しては細菌や酵母などを餌として用意する必要がある。一般的には藁やレタスなどを煮出し、その液を培地として枯草菌を増やして餌とする。枯草菌の培養が困難な場合には、市販の乾燥酵母などを利用する方法がある。
 メディアを再生する Paramecium sp. の動画。細胞口がよく分かる
メディアを再生する Paramecium sp. の動画。細胞口がよく分かる ゾウリムシは、顕微鏡下では草履(ぞうり)のような形に見える繊毛虫の1種 Paramecium caudatum の和名、広義にはゾウリムシ属 (Paramecium) に属する種を指す。単細胞生物としてはよく名を知られている。微生物自体の発見者であるオランダのレーウェンフックによって17世紀末に発見された。日本語名は、動物学者の川村多実二が1930年につけたものであり、英語名の「slipper animalcule」の「slipper」を「草履」と意訳したことに由来している。
짚신벌레는 짚신벌레과 속하는 원생생물로 대표 원생동물이다. 몸 길이 170~290µm이며 몸은 유연성이다. 앞끝은 둥글고 뒤끝은 원추형으로 뾰족하여 짚신 모양을 이룬다. 몸 전체에 균일하게 섬모가 빽빽이 자라 섬모운동을 하고, 입은 몸의 중앙부에서 깔때기 모양으로 열리며, 여기서 몸의 앞끝 왼쪽에 이르기까지 위구부(圍口部)라는 큰 홈이 계속되어 있다. 중앙부에 대핵과 소핵이 있다. 이처럼 짚신벌레는 아메바에 비해 훨씬 분화된 구조로 이루어져 있다. 담수에 살며 못·도랑·논·늪 등에 흔히 있다. 입의 뒤쪽에서 가로로 이분열하여 증식하므로, 뒤쪽 절반의 낭세포는 입을 새로 형성한다. 성숙한 개체는 접합에 의해서 유성생식을 한다. 이 경우 암수의 성별은 형태적으로는 없으나, 서로 접합 가능한 상대가 유전적으로 정해져 있으므로 수정생리학(受精生理學)이나 유전학의 연구재료로 많이 사용된다. 먹이는 짚신벌레의 한쪽에 있는 세포입이라는 구멍을 통해 안으로 들어간다. 세포입은 식도관으로 이어지고, 먹이는 식도관에서 공 모양의 식포로 원형질 속으로 들어간다. 배설물은 세포항문으로 배출한다.[1]






















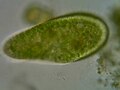
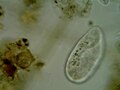
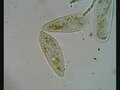


{kind=link}