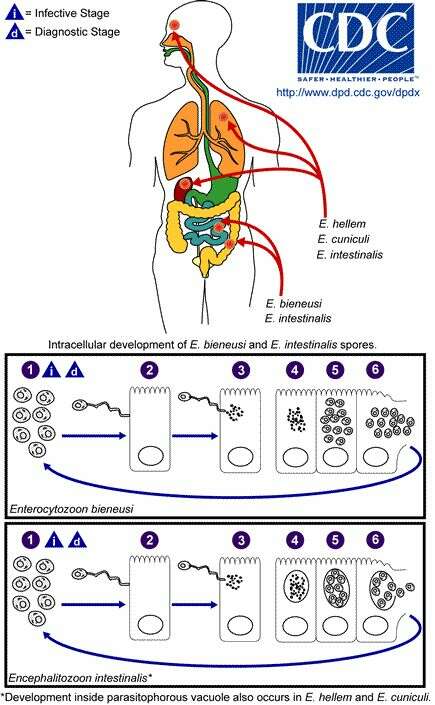
Microsporidia (Microsporidian Parasites) is a phylum of Fungi. There are 962 species of Microsporidian Parasites, in 142 genera and 32 families. It includes groups like Metchnikovellidae, Amphiacanthidae, and Buxtehudeidae.
EOL has data for 3 attributes, including: